

Atypical B cells are part of an alternative lineage of B cells that participates in responses to vaccination and infection in humans
- PMID: 33567273
- PMCID: PMC7873835
- DOI: 10.1016/j.celrep.2020.108684
Atypical B cells are part of an alternative lineage of B cells that participates in responses to vaccination and infection in humans
Abstract
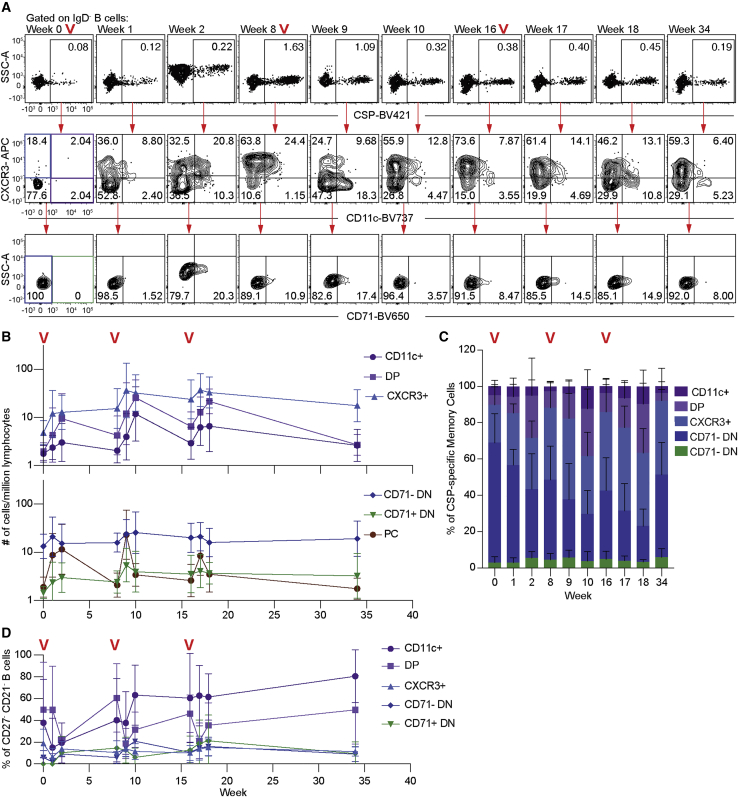
The diversity of circulating human B cells is unknown. We use single-cell RNA sequencing (RNA-seq) to examine the diversity of both antigen-specific and total B cells in healthy subjects and malaria-exposed individuals. This reveals two B cell lineages: a classical lineage of activated and resting memory B cells and an alternative lineage, which includes previously described atypical B cells. Although atypical B cells have previously been associated with disease states, the alternative lineage is common in healthy controls, as well as malaria-exposed individuals. We further track Plasmodium-specific B cells after malaria vaccination in naive volunteers. We find that alternative lineage cells are primed after the initial immunization and respond to booster doses. However, alternative lineage cells develop an atypical phenotype with repeated boosts. The data highlight that atypical cells are part of a wider alternative lineage of B cells that are a normal component of healthy immune responses.
Keywords: B cell memory; CITE-seq; alternative B cell lineage; atypical B cells; malaria; single cell RNA-seq; sporozoite; vaccination.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests S.C., N.K., B.K.L.S., and S.L.H. are salaried employees of Sanaria, the developer and owner of PfSPZ Vaccine and the investigational new drug (IND) application sponsor of the clinical trials. S.L.H. and B.K.L.S. have a financial interest in Sanaria. All other authors declare no competing interests.
Figures







Comment in
-
Atypical is the new norm, for B cells and the rest of us.Sci Immunol. 2021 Apr 2;6(58):eabi7304. doi: 10.1126/sciimmunol.abi7304. Sci Immunol. 2021. PMID: 33811062
References
-
- Andrews S.F., Chambers M.J., Schramm C.A., Plyler J., Raab J.E., Kanekiyo M., Gillespie R.A., Ransier A., Darko S., Hu J. Activation dynamics and immunoglobulin evolution of pre-existing and newly generated human memory B cell responses to influenza hemagglutinin. Immunity. 2019;51:398–410.e5. - PubMed
-
- Avery D.T., Ellyard J.I., Mackay F., Corcoran L.M., Hodgkin P.D., Tangye S.G. Increased expression of CD27 on activated human memory B cells correlates with their commitment to the plasma cell lineage. J. Immunol. 2005;174:4034–4042. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
