

Engineering adeno-associated viral vectors to evade innate immune and inflammatory responses
- PMID: 33568518
- PMCID: PMC8409505
- DOI: 10.1126/scitranslmed.abd3438
Engineering adeno-associated viral vectors to evade innate immune and inflammatory responses
Abstract
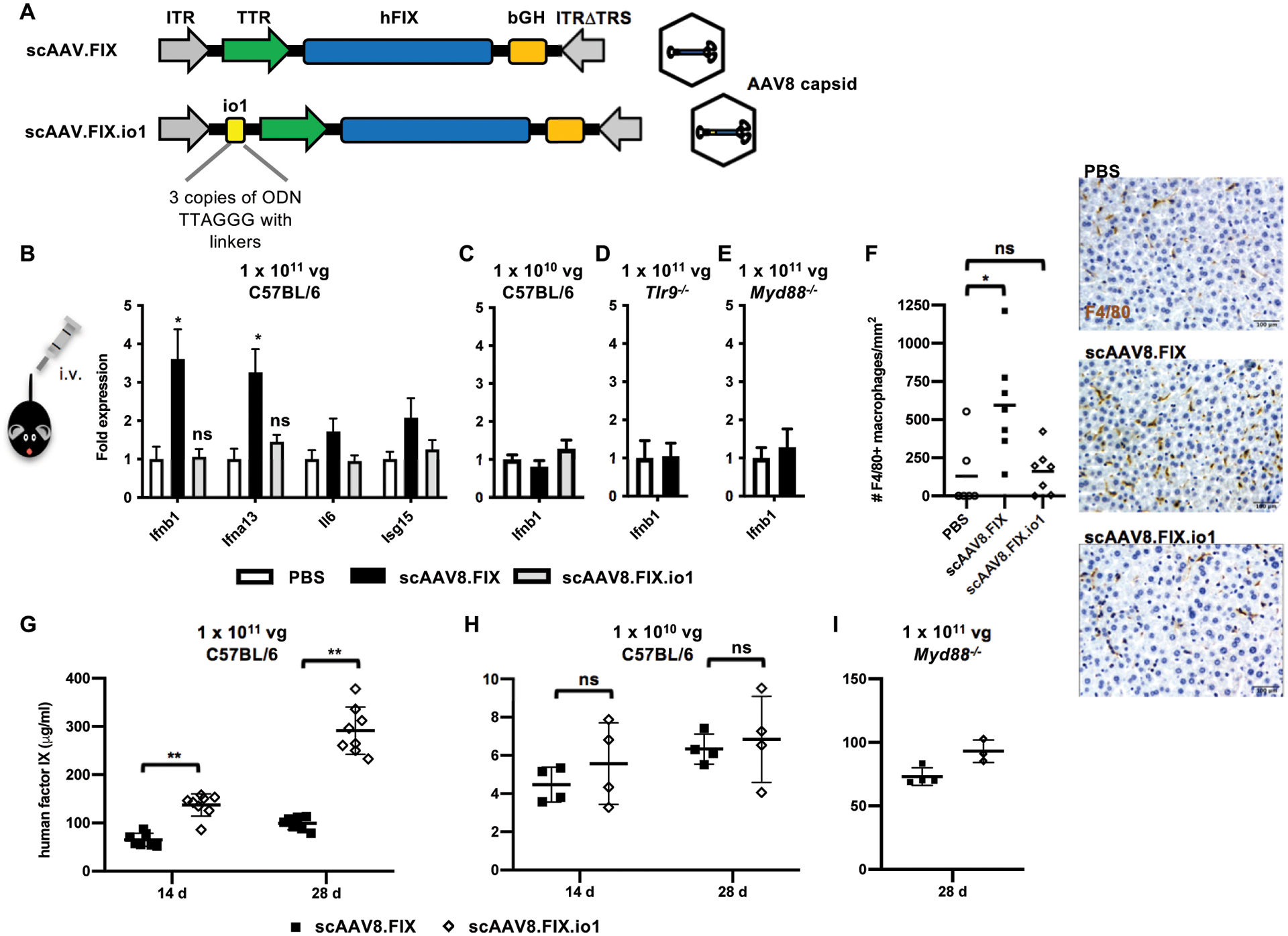
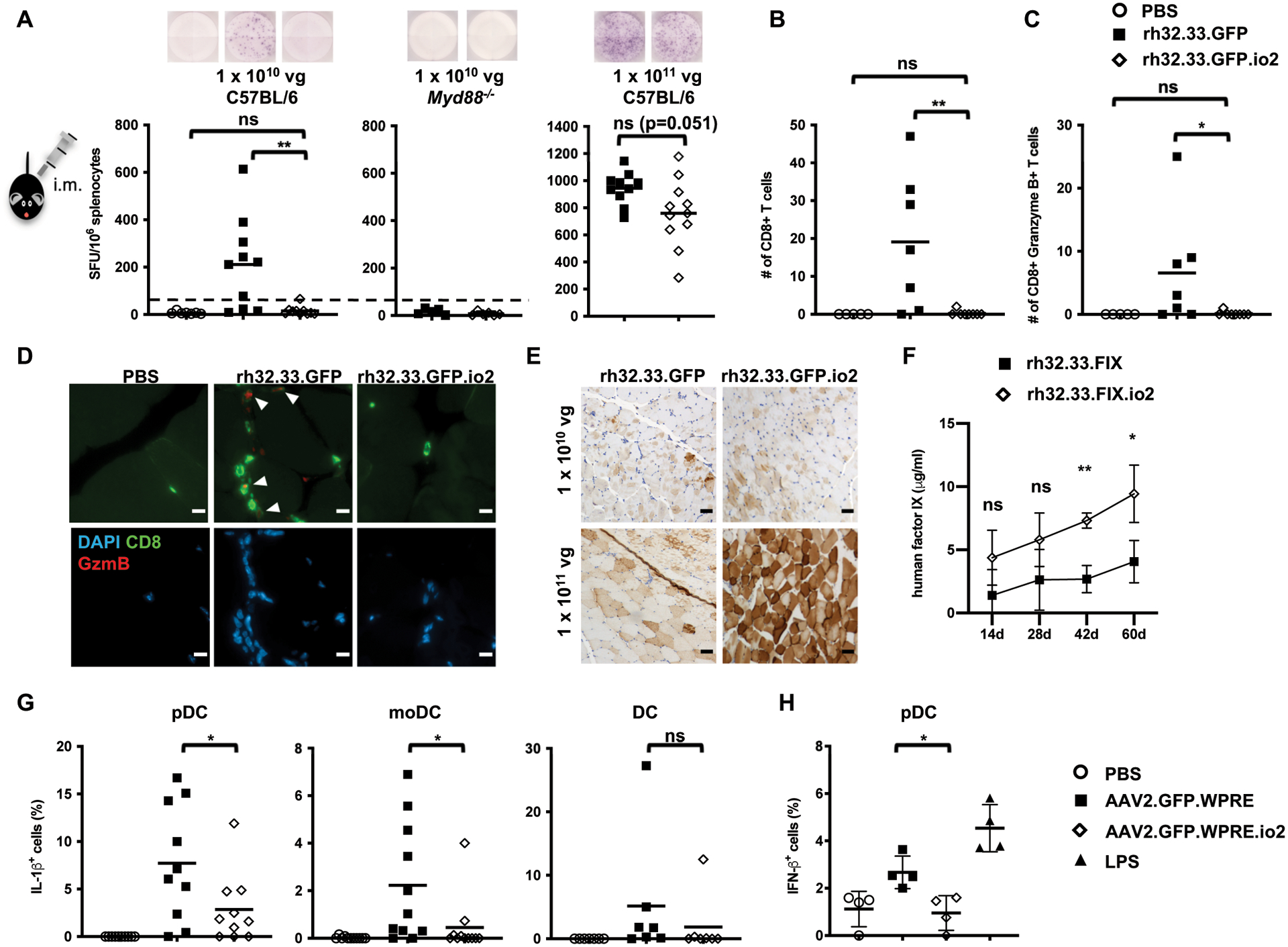
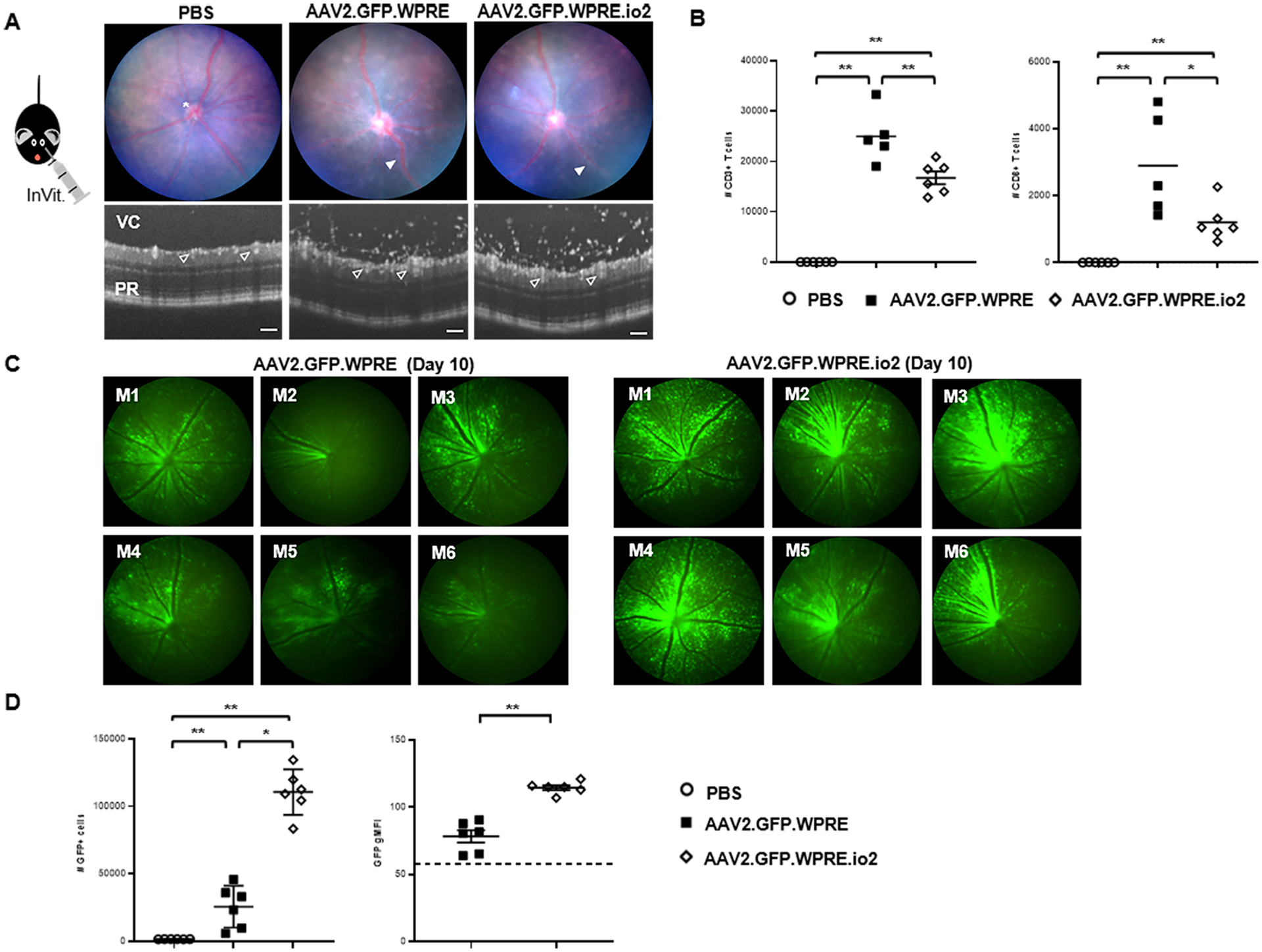
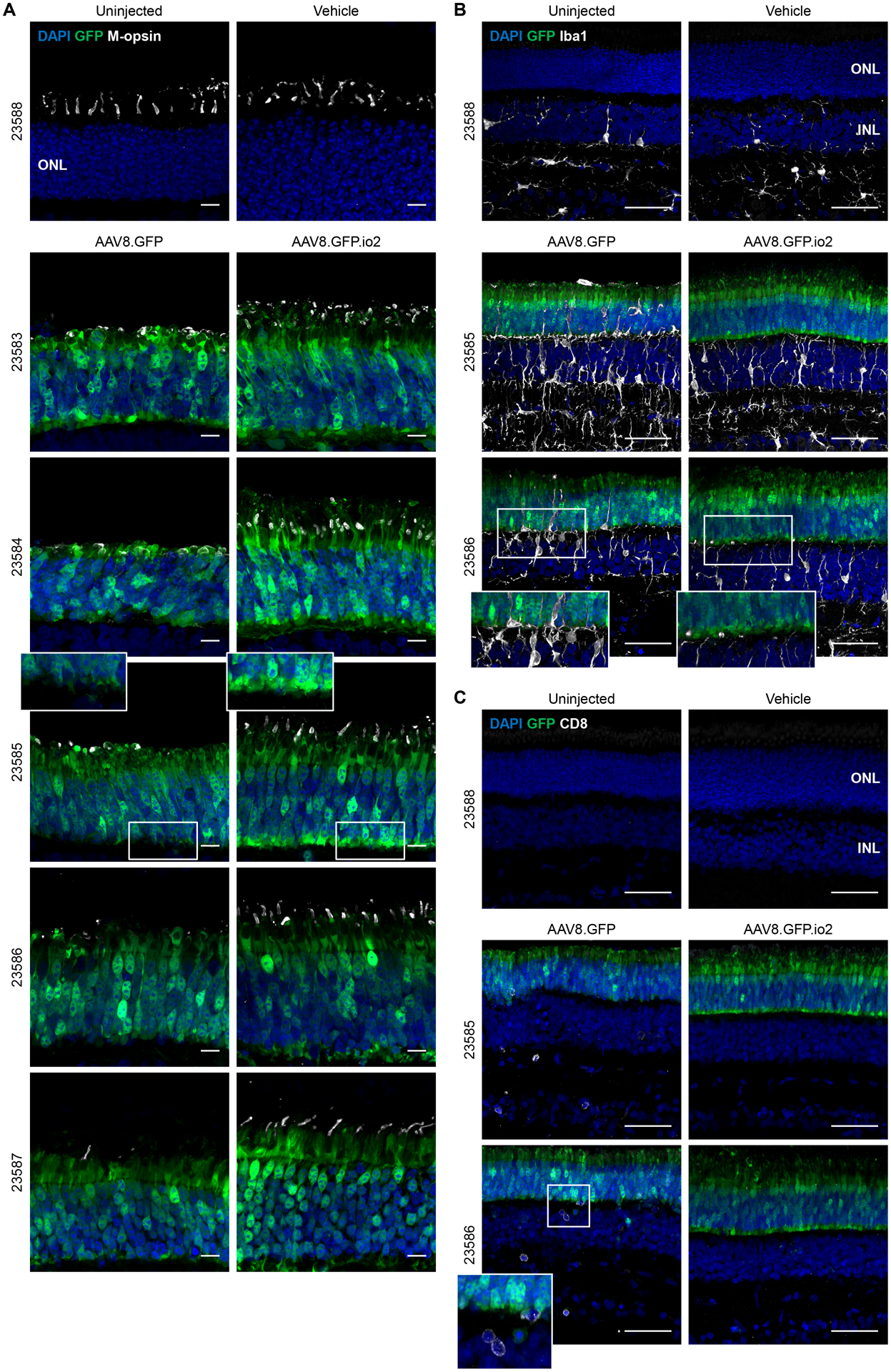
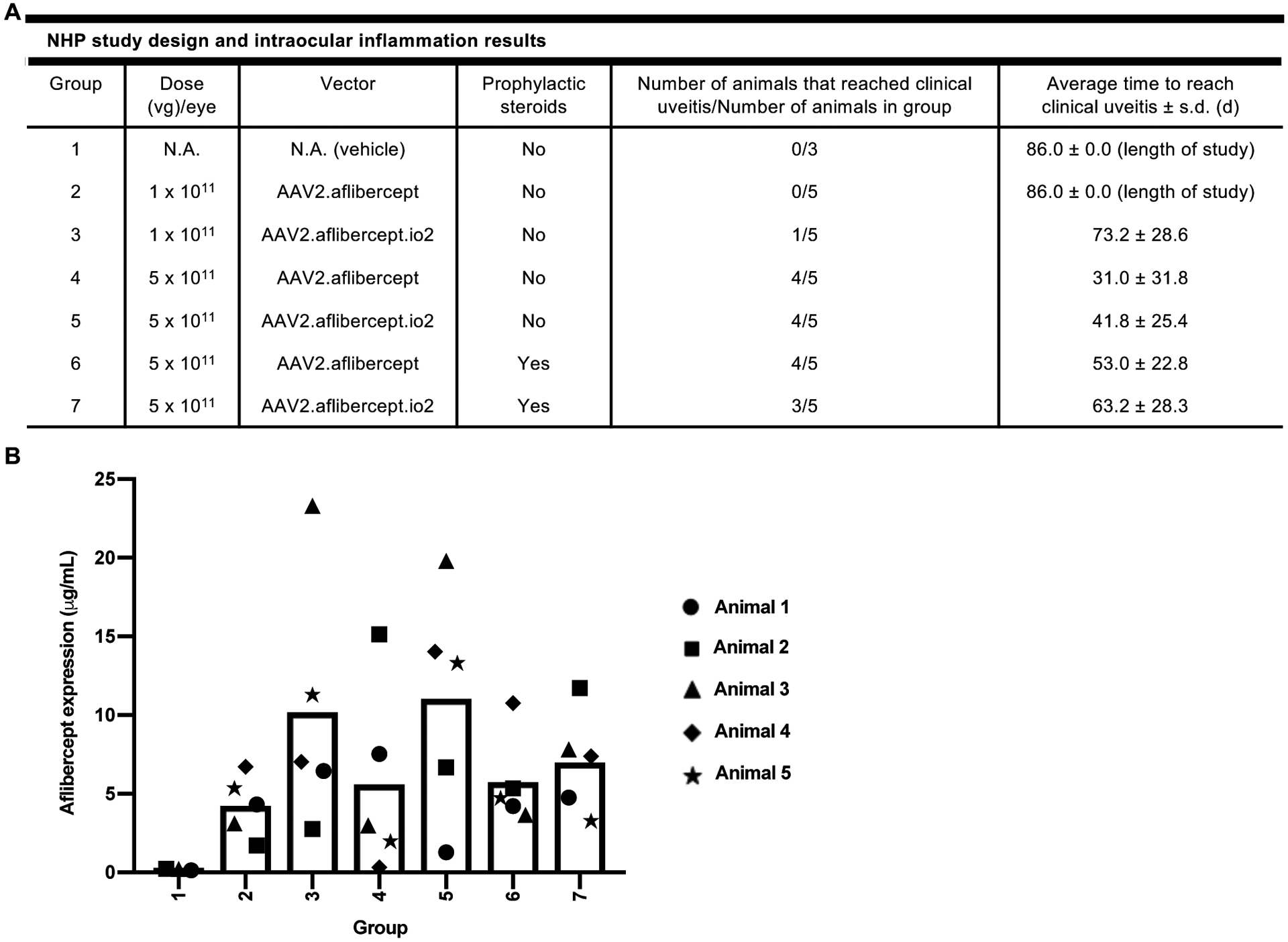
Nucleic acids are used in many therapeutic modalities, including gene therapy, but their ability to trigger host immune responses in vivo can lead to decreased safety and efficacy. In the case of adeno-associated viral (AAV) vectors, studies have shown that the genome of the vector activates Toll-like receptor 9 (TLR9), a pattern recognition receptor that senses foreign DNA. Here, we engineered AAV vectors to be intrinsically less immunogenic by incorporating short DNA oligonucleotides that antagonize TLR9 activation directly into the vector genome. The engineered vectors elicited markedly reduced innate immune and T cell responses and enhanced gene expression in clinically relevant mouse and pig models across different tissues, including liver, muscle, and retina. Subretinal administration of higher-dose AAV in pigs resulted in photoreceptor pathology with microglia and T cell infiltration. These adverse findings were avoided in the contralateral eyes of the same animals that were injected with the engineered vectors. However, intravitreal injection of higher-dose AAV in macaques, a more immunogenic route of administration, showed that the engineered vector delayed but did not prevent clinical uveitis, suggesting that other immune factors in addition to TLR9 may contribute to intraocular inflammation in this model. Our results demonstrate that linking specific immunomodulatory noncoding sequences to much longer therapeutic nucleic acids can "cloak" the vector from inducing unwanted immune responses in multiple, but not all, models. This "coupled immunomodulation" strategy may widen the therapeutic window for AAV therapies as well as other DNA-based gene transfer methods.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
-
- Hinderer C, Katz N, Buza EL, Dyer C, Goode T, Bell P, Richman LK, Wilson JM, Severe Toxicity in Nonhuman Primates and Piglets Following High-Dose Intravenous Administration of an Adeno-Associated Virus Vector Expressing Human SMN. Hum Gene Ther 29, 285–298 (2018); published online EpubMar (10.1089/hum.2018.015). - DOI - PMC - PubMed
-
- Mendell JR, Al-Zaidy S, Shell R, Arnold WD, Rodino-Klapac LR, Prior TW, Lowes L, Alfano L, Berry K, Church K, Kissel JT, Nagendran S, L’Italien J, Sproule DM, Wells C, Cardenas JA, Heitzer MD, Kaspar A, Corcoran S, Braun L, Likhite S, Miranda C, Meyer K, Foust KD, Burghes AHM, Kaspar BK, Single-Dose Gene-Replacement Therapy for Spinal Muscular Atrophy. N Engl J Med 377, 1713–1722 (2017); published online EpubNov 2 (10.1056/NEJMoa1706198). - DOI - PubMed
-
- Cukras C, Wiley HE, Jeffrey BG, Sen HN, Turriff A, Zeng Y, Vijayasarathy C, Marangoni D, Ziccardi L, Kjellstrom S, Park TK, Hiriyanna S, Wright JF, Colosi P, Wu Z, Bush RA, Wei LL, Sieving PA, Retinal AAV8-RS1 Gene Therapy for X-Linked Retinoschisis: Initial Findings from a Phase I/IIa Trial by Intravitreal Delivery. Mol Ther 26, 2282–2294 (2018); published online EpubSep 5 (10.1016/j.ymthe.2018.05.025). - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
