

Loss of TGFβ signaling increases alternative end-joining DNA repair that sensitizes to genotoxic therapies across cancer types
- PMID: 33568520
- PMCID: PMC8208885
- DOI: 10.1126/scitranslmed.abc4465
Loss of TGFβ signaling increases alternative end-joining DNA repair that sensitizes to genotoxic therapies across cancer types
Abstract
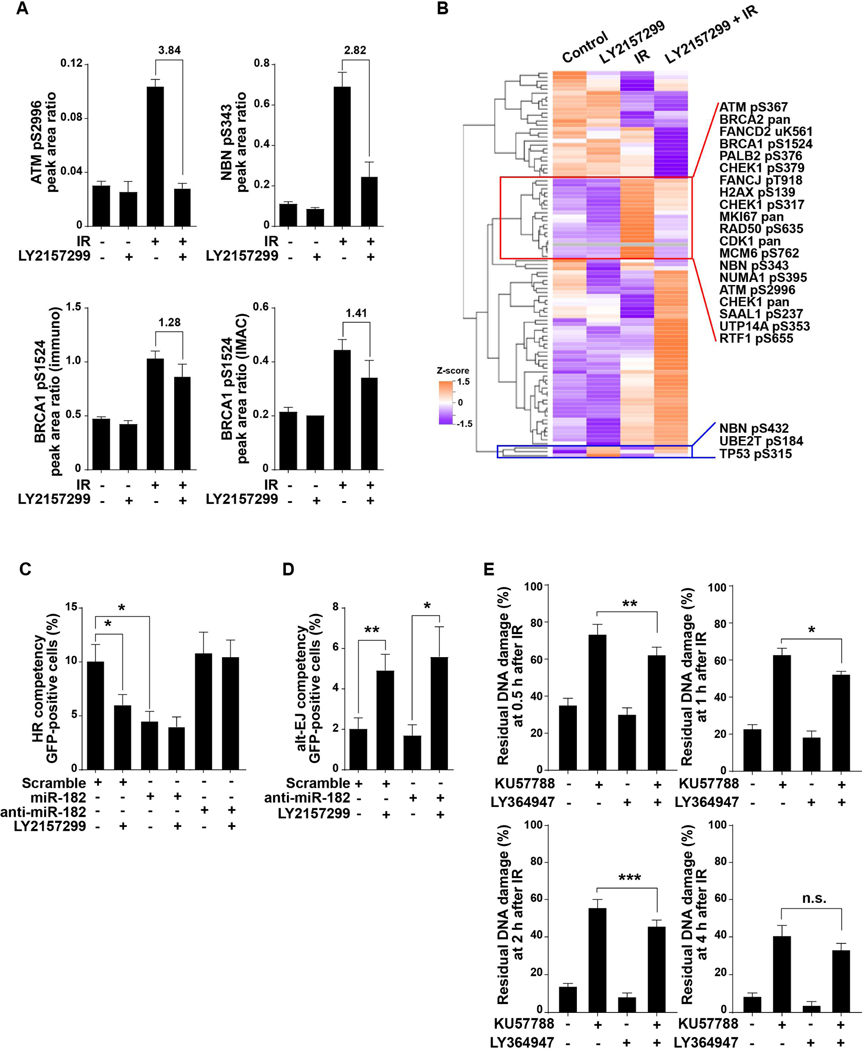
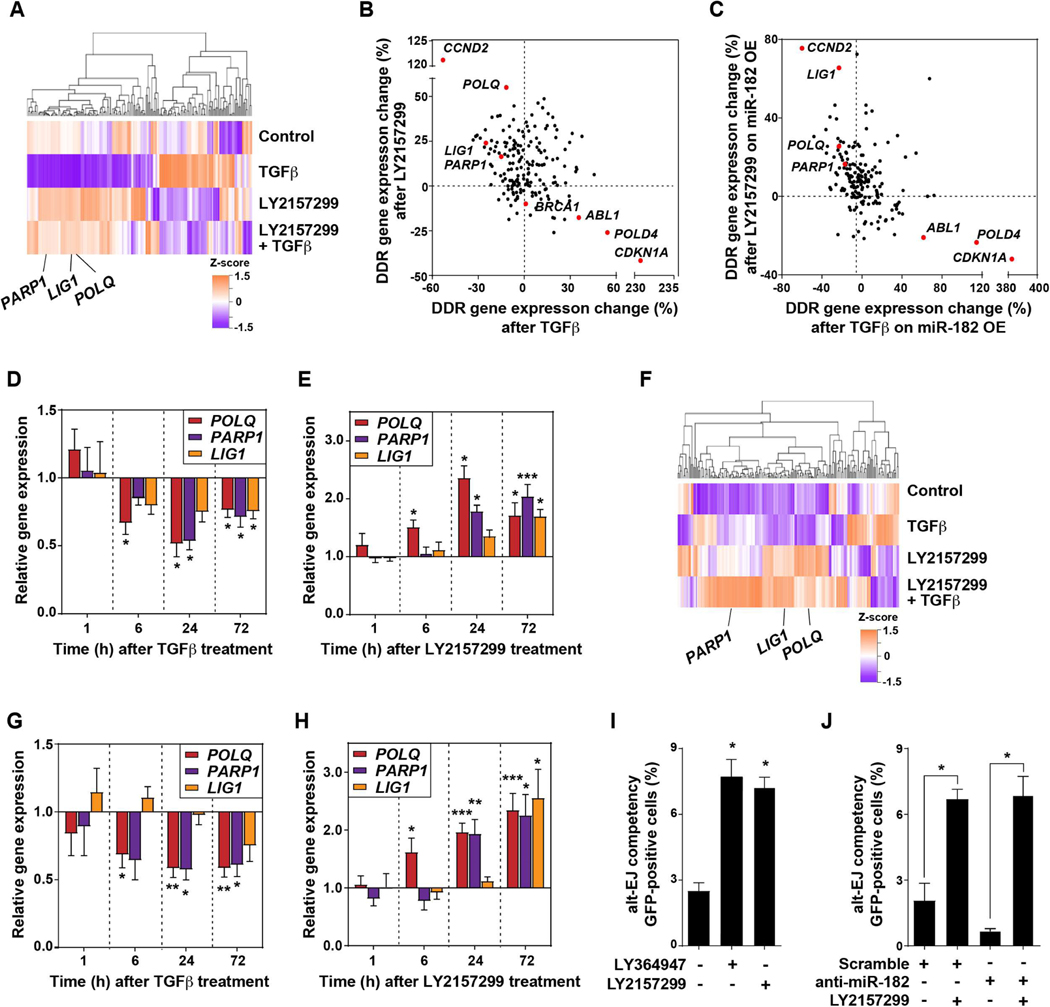
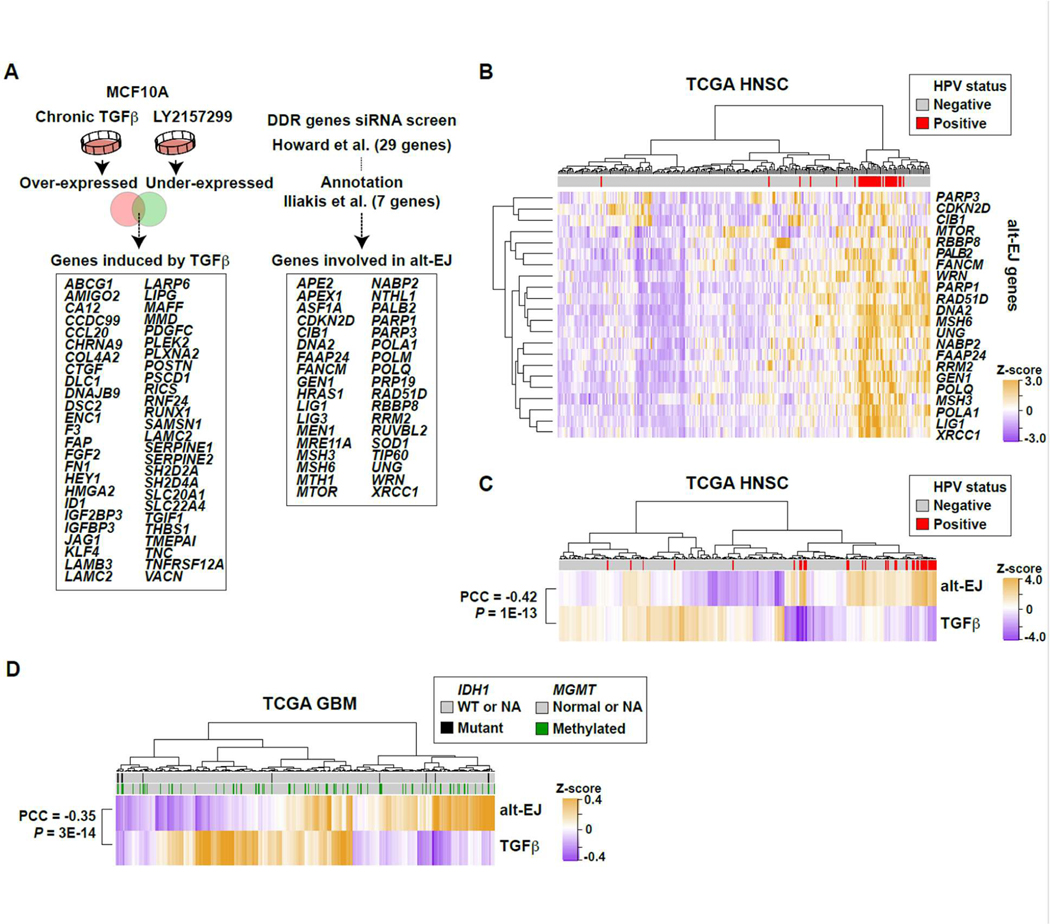
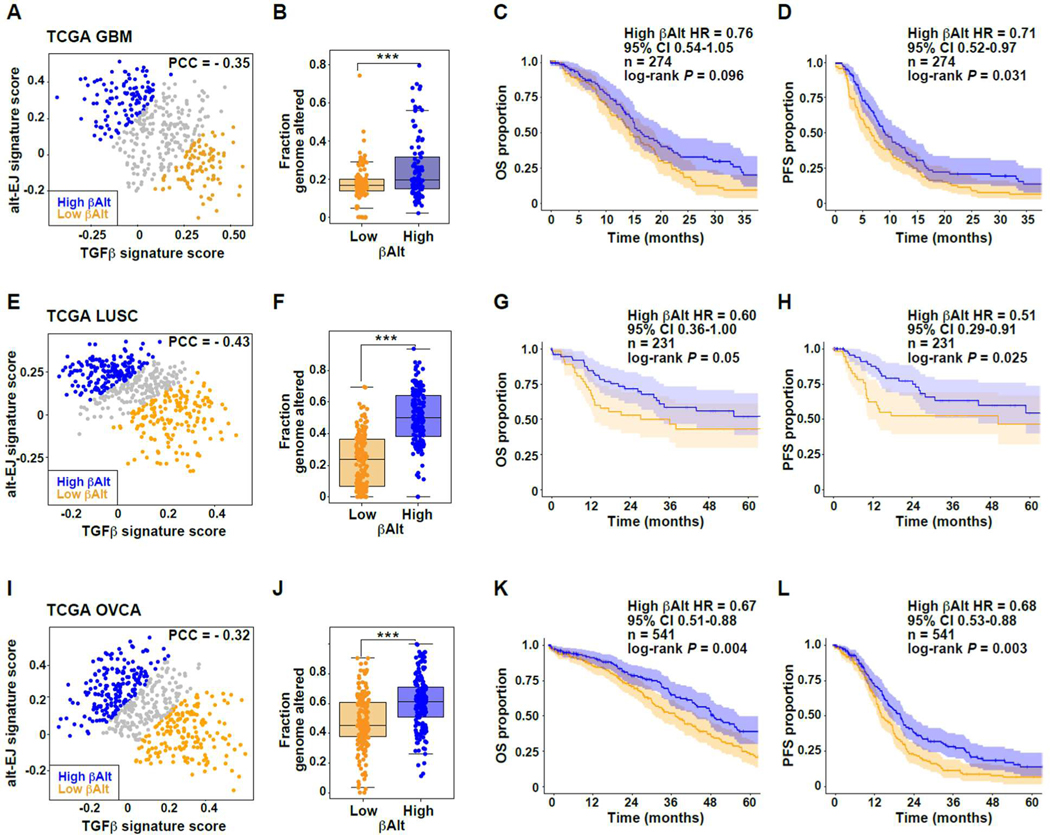
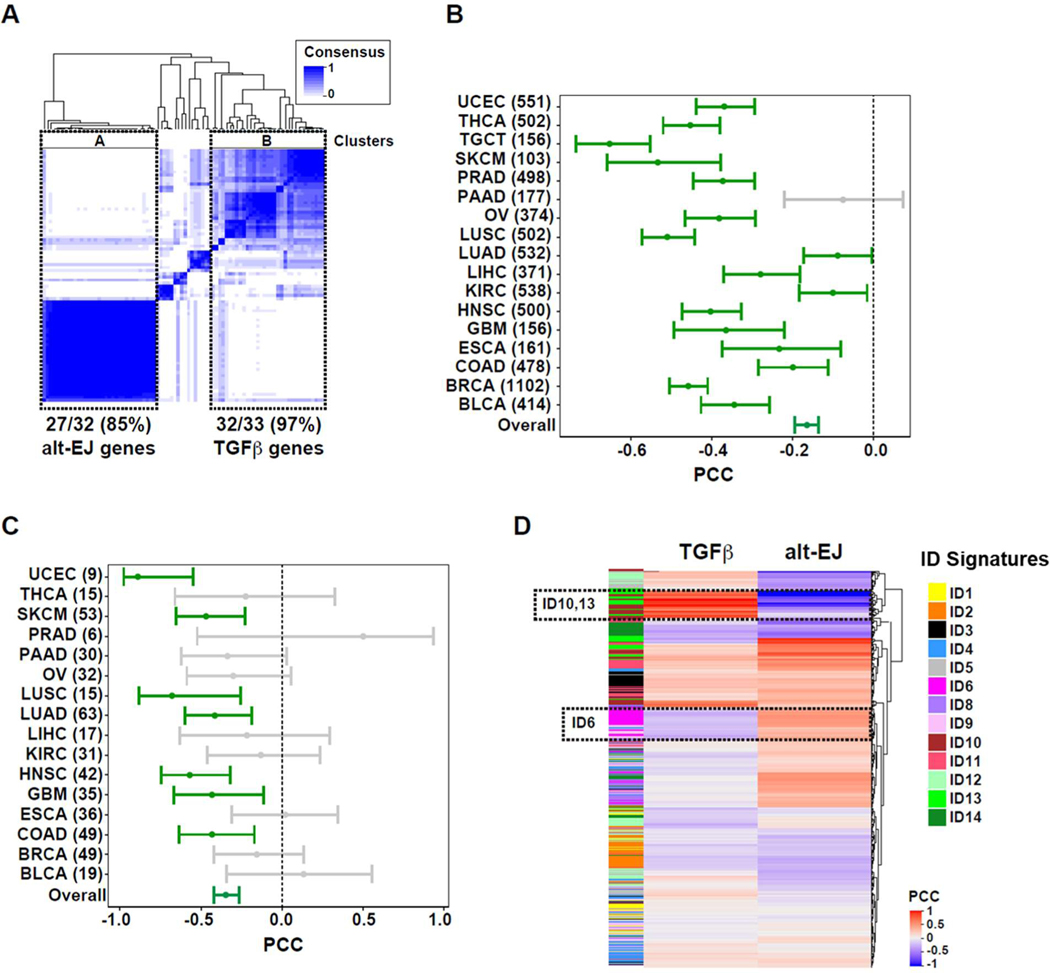
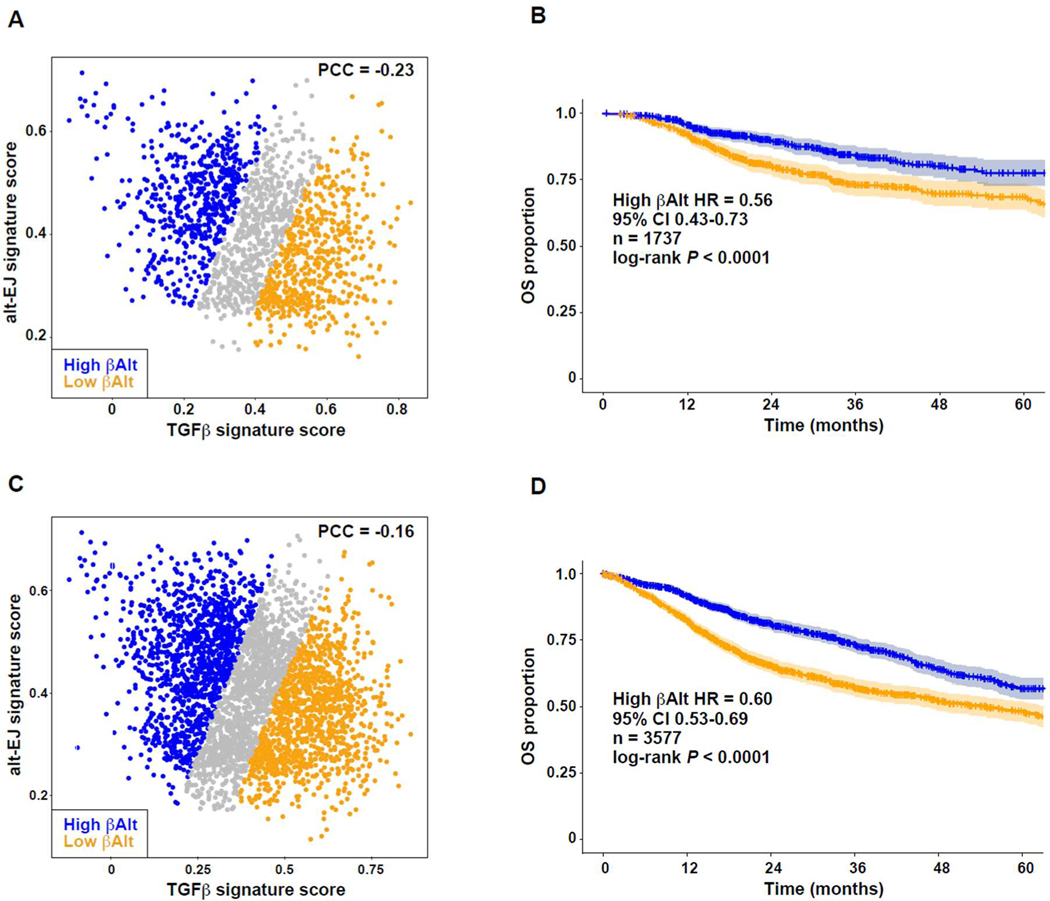
Among the pleotropic roles of transforming growth factor-β (TGFβ) signaling in cancer, its impact on genomic stability is least understood. Inhibition of TGFβ signaling increases use of alternative end joining (alt-EJ), an error-prone DNA repair process that typically functions as a "backup" pathway if double-strand break repair by homologous recombination or nonhomologous end joining is compromised. However, the consequences of this functional relationship on therapeutic vulnerability in human cancer remain unknown. Here, we show that TGFβ broadly controls the DNA damage response and suppresses alt-EJ genes that are associated with genomic instability. Mechanistically based TGFβ and alt-EJ gene expression signatures were anticorrelated in glioblastoma, squamous cell lung cancer, and serous ovarian cancer. Consistent with error-prone repair, more of the genome was altered in tumors classified as low TGFβ and high alt-EJ, and the corresponding patients had better outcomes. Pan-cancer analysis of solid neoplasms revealed that alt-EJ genes were coordinately expressed and anticorrelated with TGFβ competency in 16 of 17 cancer types tested. Moreover, regardless of cancer type, tumors classified as low TGFβ and high alt-EJ were characterized by an insertion-deletion mutation signature containing short microhomologies and were more sensitive to genotoxic therapy. Collectively, experimental studies revealed that loss or inhibition of TGFβ signaling compromises the DNA damage response, resulting in ineffective repair by alt-EJ. Translation of this mechanistic relationship into gene expression signatures identified a robust anticorrelation that predicts response to genotoxic therapies, thereby expanding the potential therapeutic scope of TGFβ biology.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
References
-
- Korkut A, Zaidi S, Kanchi RS, Rao S, Gough NR, Schultz A, Li X, Lorenzi PL, Berger AC, Robertson G, Kwong LN, Datto M, Roszik J, Ling S, Ravikumar V, Manyam G, Rao A, Shelley S, Liu Y, Ju Z, Hansel D, de Velasco G, Pennathur A, Andersen JB, O’Rourke CJ, Ohshiro K, Jogunoori W, Nguyen BN, Li S, Osmanbeyoglu HU, Ajani JA, Mani SA, Houseman A, Wiznerowicz M, Chen J, Gu S, Ma W, Zhang J, Tong P, Cherniack AD, Deng C, Resar L, Weinstein JN, Mishra L, Akbani R, A pan-cancer analysis reveals high-frequency genetic alterations in mediators of signaling by the TGF-beta superfamily. Cell Syst. 7, 422–437.e427 (2018). - PMC - PubMed
-
- Glick AB, Weinberg WC, Wu IH, Quan W, Yuspa SH, Transforming growth factor β1 suppresses genomic instability independent of a G1 arrest, p53, and Rb. Cancer Res. 56, 3645–3650 (1996). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
