

miR-16-5p Promotes Erythroid Maturation of Erythroleukemia Cells by Regulating Ribosome Biogenesis
- PMID: 33572085
- PMCID: PMC7915806
- DOI: 10.3390/ph14020137
miR-16-5p Promotes Erythroid Maturation of Erythroleukemia Cells by Regulating Ribosome Biogenesis
Abstract
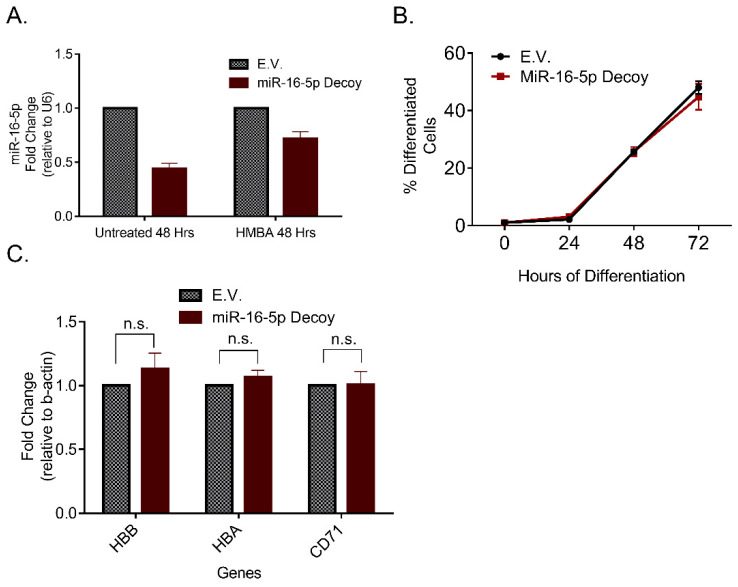
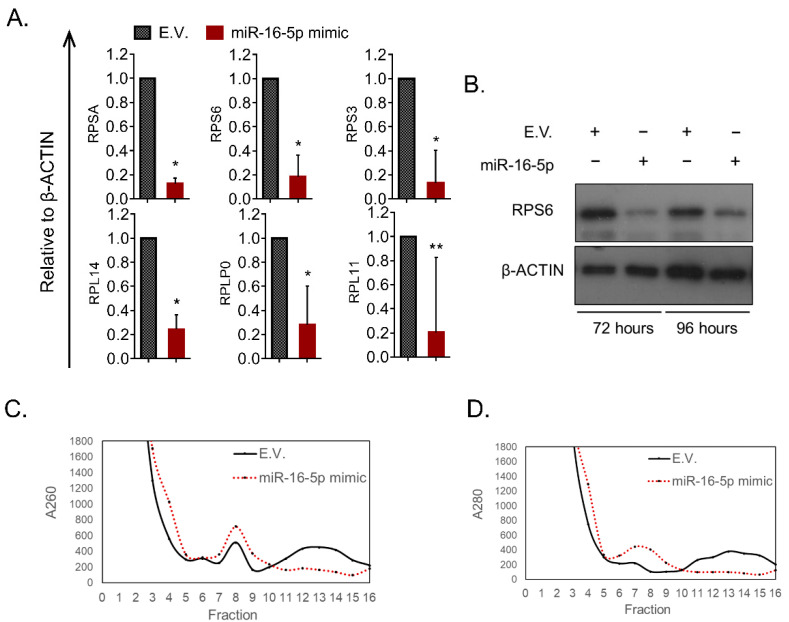
miRNAs constitute a class of non-coding RNA that act as powerful epigenetic regulators in animal and plant cells. In order to identify putative tumor-suppressor miRNAs we profiled the expression of various miRNAs during differentiation of erythroleukemia cells. RNA was purified before and after differentiation induction and subjected to quantitative RT-PCR. The majority of the miRNAs tested were found upregulated in differentiated cells with miR-16-5p showing the most significant increase. Functional studies using gain- and loss-of-function constructs proposed that miR-16-5p has a role in promoting the erythroid differentiation program of murine erythroleukemia (MEL) cells. In order to identify the underlying mechanism of action, we utilized bioinformatic in-silico platforms that incorporate predictions for the genes targeted by miR-16-5p. Interestingly, ribosome constituents, as well as ribosome biogenesis factors, were overrepresented among the miR-16-5p predicted gene targets. Accordingly, biochemical experiments showed that, indeed, miR-16-5p could modulate the levels of independent ribosomal proteins, and the overall ribosomal levels in cultured cells. In conclusion, miR-16-5p is identified as a differentiation-promoting agent in erythroleukemia cells, demonstrating antiproliferative activity, likely as a result of its ability to target the ribosomal machinery and restore any imbalanced activity imposed by the malignancy and the blockade of differentiation.
Keywords: cancer; erythroid differentiation; erythroleukemia; miR-16-5p; miRNA therapeutics; ribosomes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
LinkOut - more resources
Full Text Sources
Other Literature Sources
