

SOX2 is required independently in both stem and differentiated cells for pituitary tumorigenesis in p27-null mice
- PMID: 33574062
- PMCID: PMC7896314
- DOI: 10.1073/pnas.2017115118
SOX2 is required independently in both stem and differentiated cells for pituitary tumorigenesis in p27-null mice
Abstract
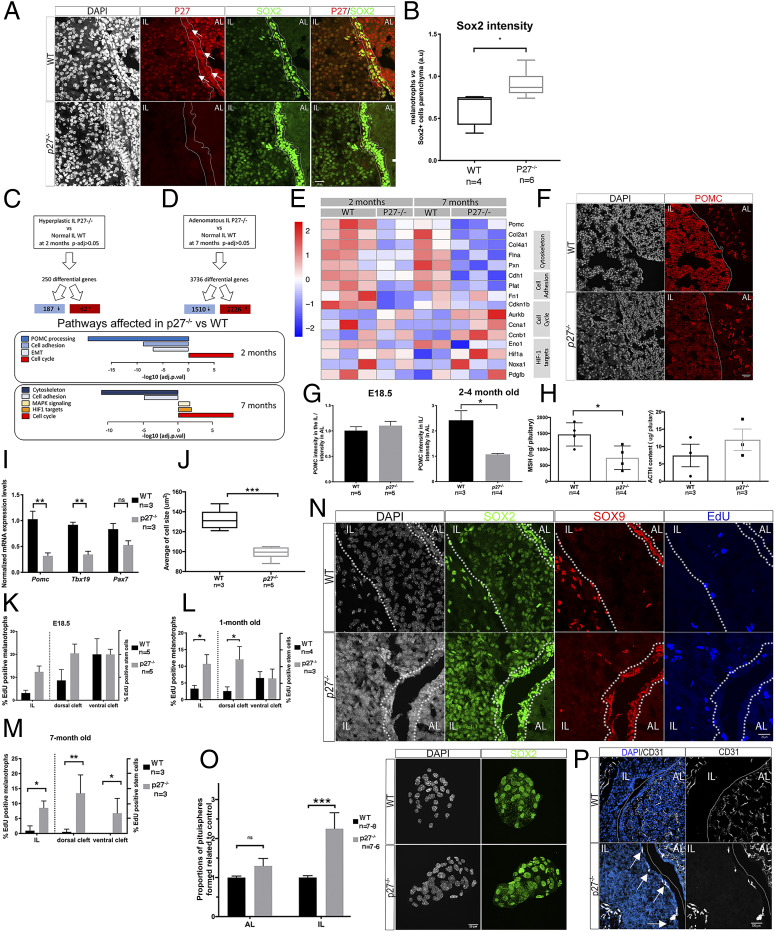
P27, a cell cycle inhibitor, is also able to drive repression of Sox2 This interaction plays a crucial role during development of p27-/- pituitary tumors because loss of one copy of Sox2 impairs tumorigenesis [H. Li et al., Cell Stem Cell 11, 845-852 (2012)]. However, SOX2 is expressed in both endocrine and stem cells (SCs), and its contribution to tumorigenesis in either cell type is unknown. We have thus explored the cellular origin and mechanisms underlying endocrine tumorigenesis in p27-/- pituitaries. We found that pituitary hyperplasia is associated with reduced cellular differentiation, in parallel with increased levels of SOX2 in stem and endocrine cells. Using conditional loss-of-function and lineage tracing approaches, we show that SOX2 is required cell autonomously in p27-/- endocrine cells for these to give rise to tumors, and in SCs for promotion of tumorigenesis. This is supported by studies deleting the Sox2 regulatory region 2 (Srr2), the target of P27 repressive action. Single cell transcriptomic analysis further reveals that activation of a SOX2-dependent MAPK pathway in SCs is important for tumorigenesis. Altogether, our data highlight different aspects of the role of SOX2 following loss of p27, according to cellular context, and uncover an unexpected SOX2-dependent tumor-promoting role for SCs. Our results imply that targeting SCs, in addition to tumor cells, may represent an efficient antitumoral strategy in certain contexts.
Keywords: pituitary; stem cell; tumorigenesis.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures






References
-
- Andoniadou C. L., et al. , Sox2(+) stem/progenitor cells in the adult mouse pituitary support organ homeostasis and have tumor-inducing potential. Cell Stem Cell 13, 433–445 (2013). - PubMed
-
- Lamolet B., et al. , A pituitary cell-restricted T box factor, Tpit, activates POMC transcription in cooperation with Pitx homeoproteins. Cell 104, 849–859 (2001). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
