

Rare genetic variants affecting urine metabolite levels link population variation to inborn errors of metabolism
- PMID: 33574263
- PMCID: PMC7878905
- DOI: 10.1038/s41467-020-20877-8
Rare genetic variants affecting urine metabolite levels link population variation to inborn errors of metabolism
Erratum in
-
Author Correction: Rare genetic variants affecting urine metabolite levels link population variation to inborn errors of metabolism.Nat Commun. 2021 Oct 6;12(1):5938. doi: 10.1038/s41467-021-26242-7. Nat Commun. 2021. PMID: 34615878 Free PMC article. No abstract available.
Abstract
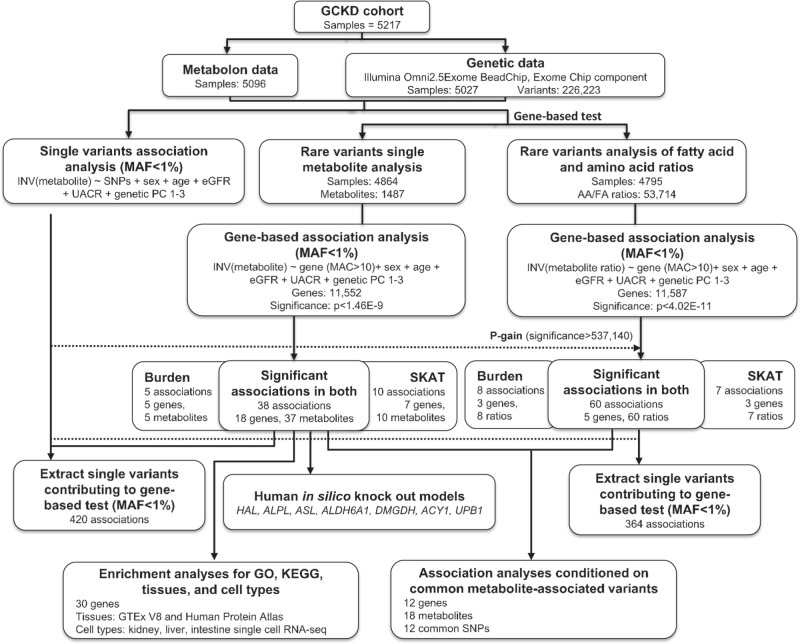
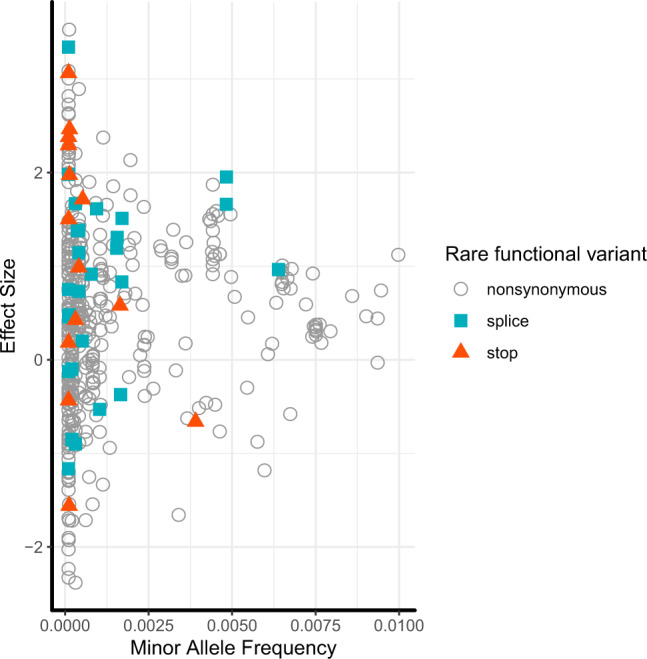
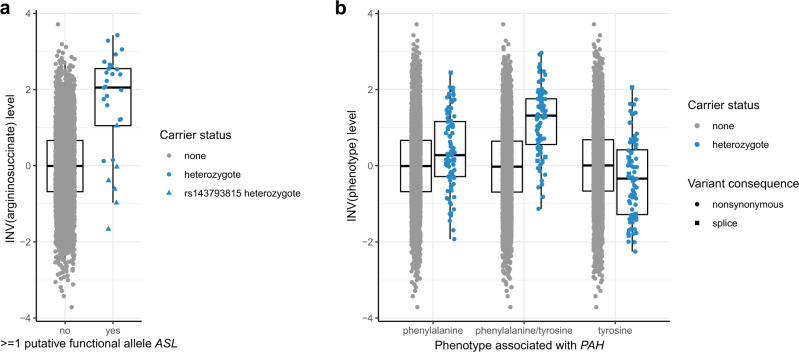
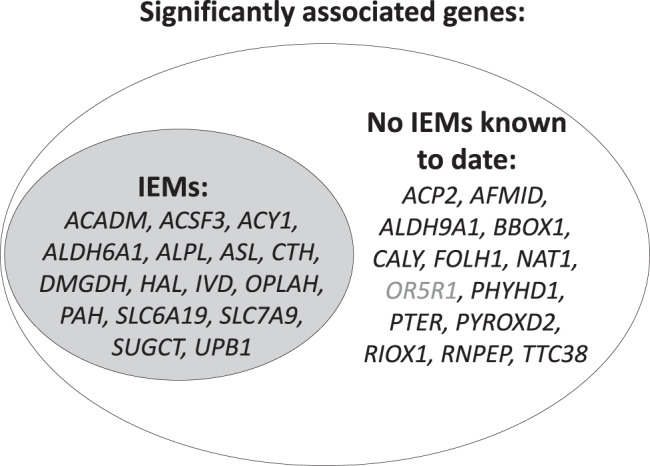
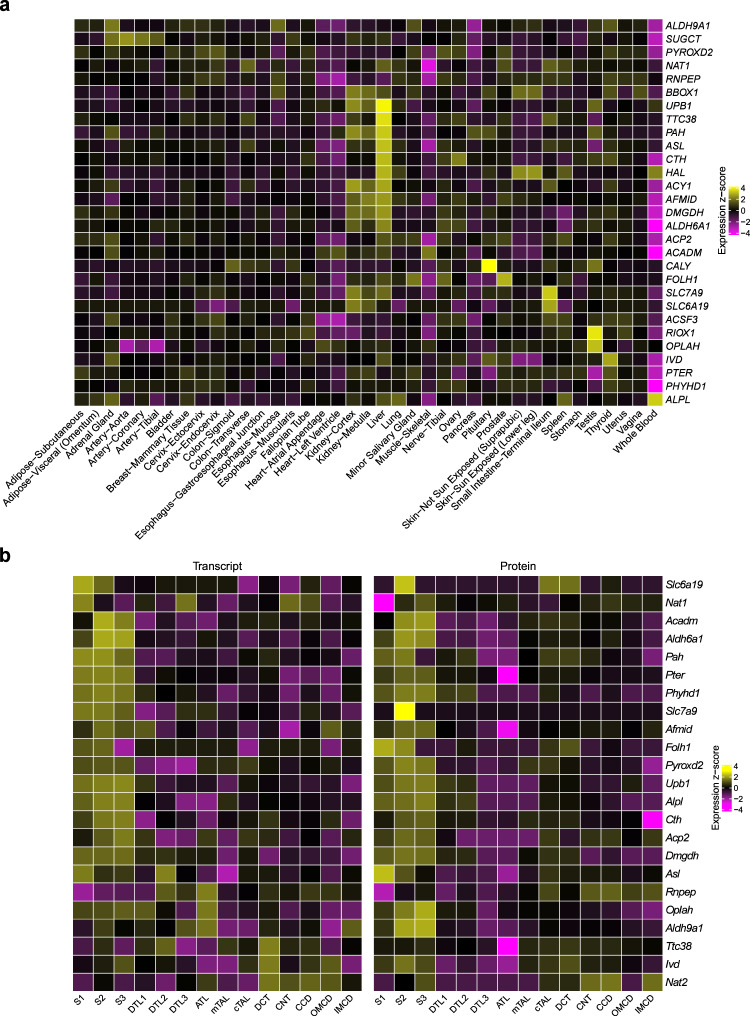
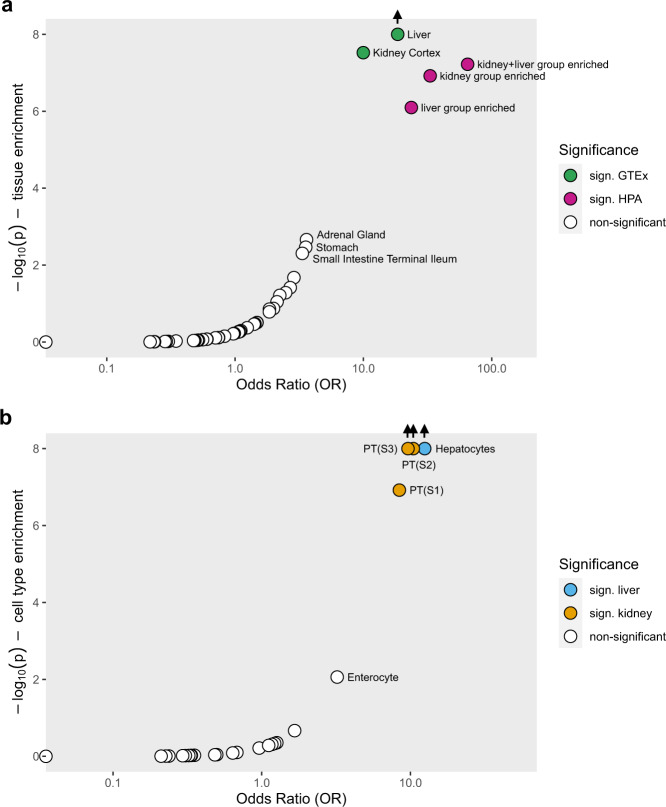
Metabolite levels in urine may provide insights into genetic mechanisms shaping their related pathways. We therefore investigate the cumulative contribution of rare, exonic genetic variants on urine levels of 1487 metabolites and 53,714 metabolite ratios among 4864 GCKD study participants. Here we report the detection of 128 significant associations involving 30 unique genes, 16 of which are known to underlie inborn errors of metabolism. The 30 genes are strongly enriched for shared expression in liver and kidney (odds ratio = 65, p-FDR = 3e-7), with hepatocytes and proximal tubule cells as driving cell types. Use of UK Biobank whole-exome sequencing data links genes to diseases connected to the identified metabolites. In silico constraint-based modeling of gene knockouts in a virtual whole-body, organ-resolved metabolic human correctly predicts the observed direction of metabolite changes, highlighting the potential of linking population genetics to modeling. Our study implicates candidate variants and genes for inborn errors of metabolism.
Conflict of interest statement
J.M. and D.F.F. are full time employees of Bayer AG, Division Pharma. All remaining authors declare no competing interests.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
