

Ornithine decarboxylase genes contribute to S-RNase-independent pollen rejection
- PMID: 33576789
- PMCID: PMC8154068
- DOI: 10.1093/plphys/kiab062
Ornithine decarboxylase genes contribute to S-RNase-independent pollen rejection
Abstract
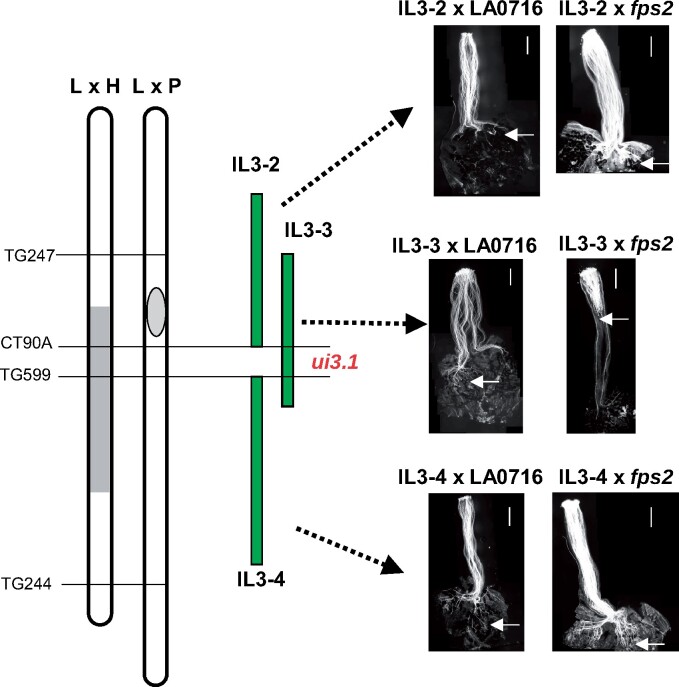
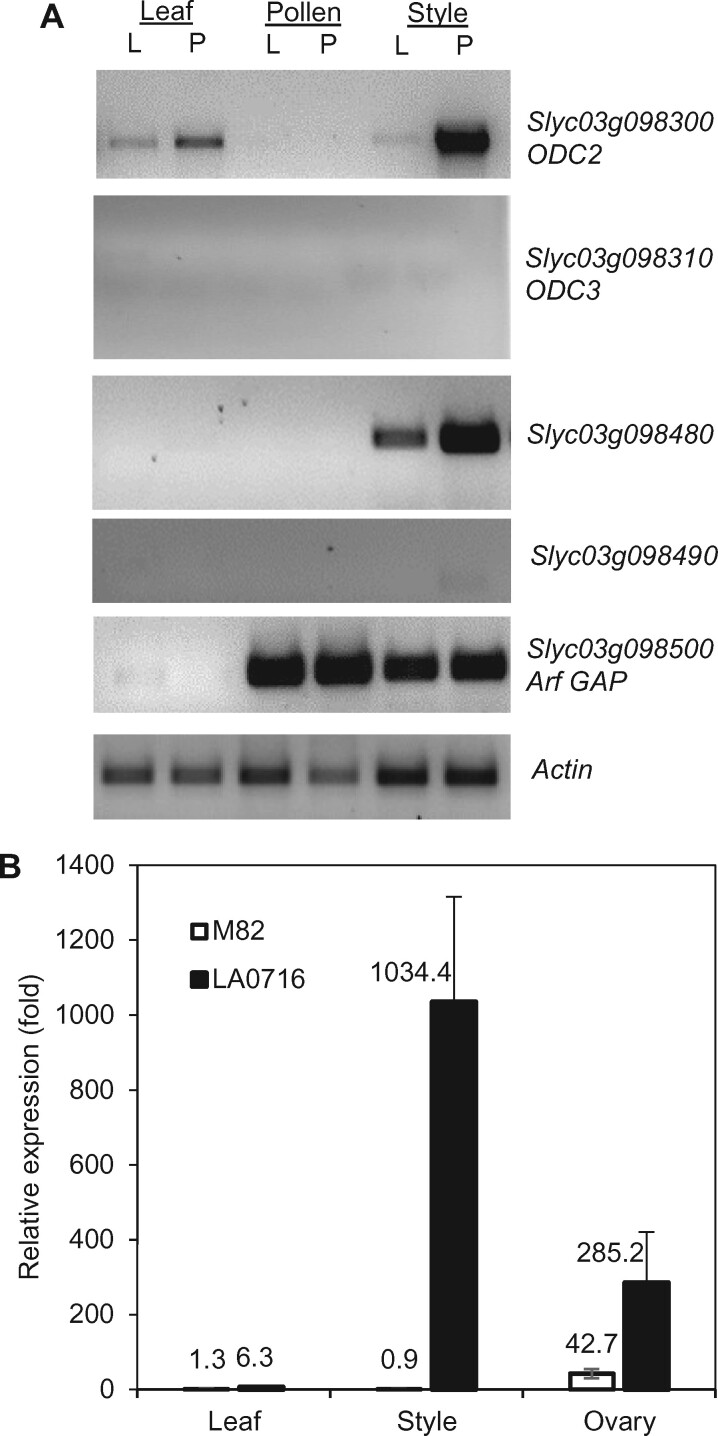
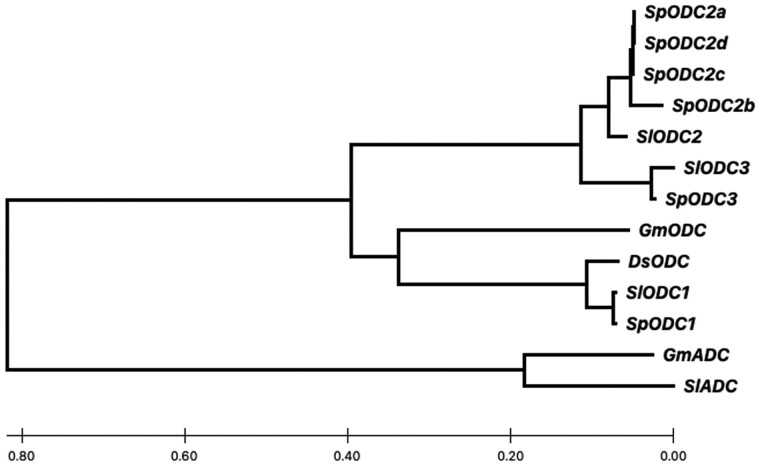
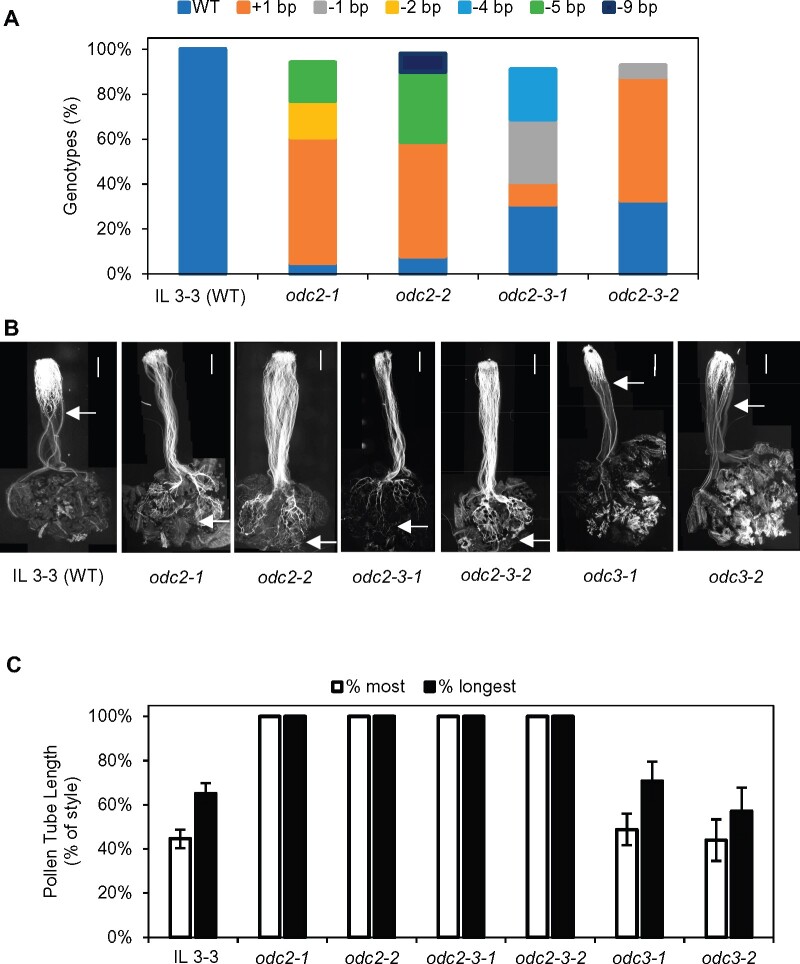
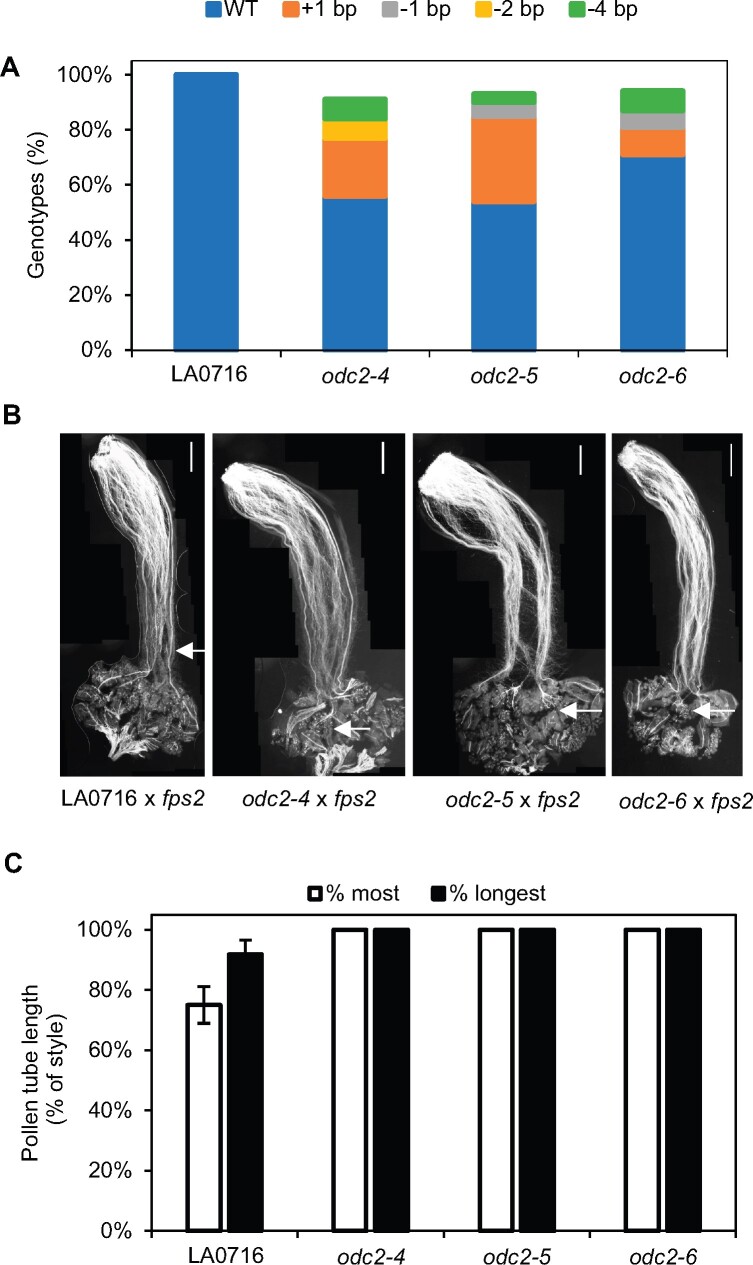
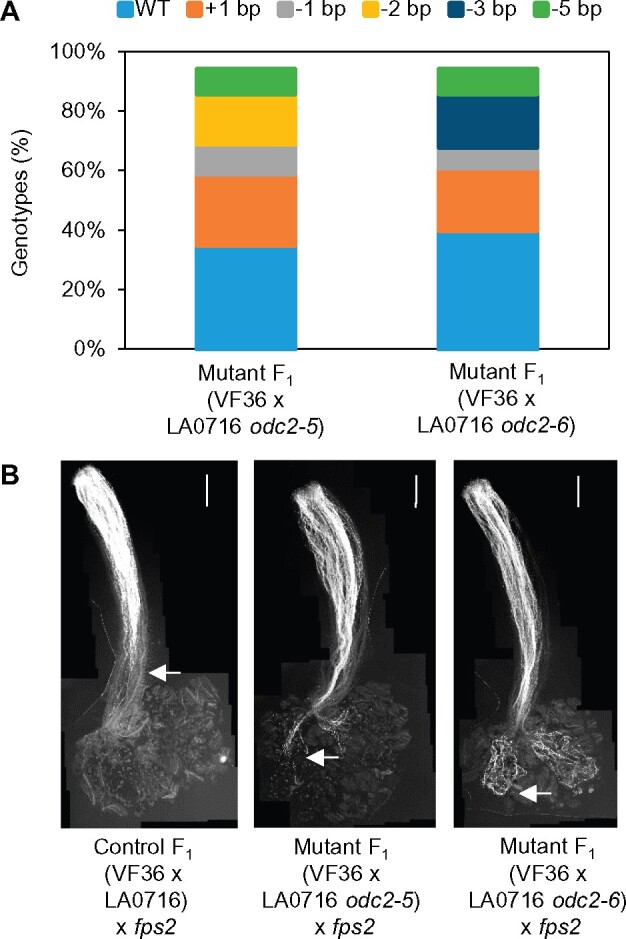
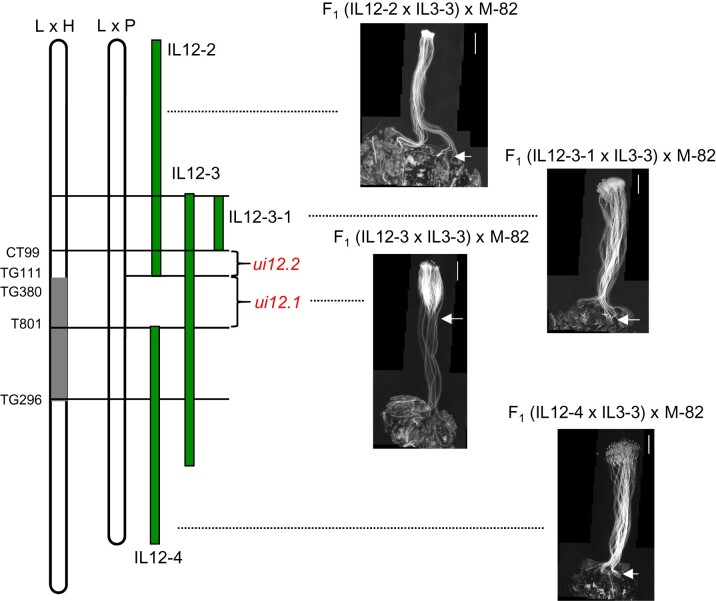
Unilateral incompatibility (UI) manifests as pollen rejection in the pistil, typically when self-incompatible (SI) species are pollinated by self-compatible (SC) relatives. In the Solanaceae, UI occurs when pollen lack resistance to stylar S-RNases, but other, S-RNase-independent mechanisms exist. Pistils of the wild tomato Solanum pennellii LA0716 (SC) lack S-RNase yet reject cultivated tomato (Solanum lycopersicum, SC) pollen. In this cross, UI results from low pollen expression of a farnesyl pyrophosphate synthase gene (FPS2) in S. lycopersicum. Using pollen from fps2-/- loss-of-function mutants in S. pennellii, we identified a pistil factor locus, ui3.1, required for FPS2-based pollen rejection. We mapped ui3.1 to an interval containing 108 genes situated on the IL 3-3 introgression. This region includes a cluster of ornithine decarboxylase (ODC2) genes, with four copies in S. pennellii, versus one in S. lycopersicum. Expression of ODC2 transcript was 1,034-fold higher in S. pennellii than in S. lycopersicum styles. Pistils of odc2-/- knockout mutants in IL 3-3 or S. pennellii fail to reject fps2 pollen and abolish transmission ratio distortion (TRD) associated with FPS2. Pollen of S. lycopersicum express low levels of FPS2 and are compatible on IL 3-3 pistils, but incompatible on IL 12-3 × IL 3-3 hybrids, which express both ODC2 and ui12.1, a locus thought to encode the SI proteins HT-A and HT-B. TRD observed in F2 IL 12-3 × IL 3-3 points to additional ODC2-interacting pollen factors on both chromosomes. Thus, ODC2 genes contribute to S-RNase independent UI and interact genetically with ui12.1 to strengthen pollen rejection.
© American Society of Plant Biologists 2021. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures

References
-
- Aloisi I, Distefano G, Antognoni F, Potente G, Parrotta L, Faleri C, Gentile A, Bennici S, Mareri L, Cai G, Del Duca S (2020) Temperature-dependent compatible and incompatible pollen-style interactions in Citrus clementina Hort. ex Tan. Show different transglutaminase features and polyamine pattern. Front Plant Sci 11: 1018. - PMC - PubMed
-
- Alseekh S, Ofner I, Pleban T, Tripodi P, Di Dato F, Cammareri M, Mohammad A, Grandillo S, Fernie AR, Zamir D (2013) Resolution by recombination: breaking up Solanum pennellii introgressions. Trends Plant Sci 18: 536–538 - PubMed
-
- Baek YS, Covey PA, Petersen JJ, Chetelat RT, McClure B, Bedinger P (2015) Testing the ‘SI × SC rule’: pollen–pistil interactions in interspecific crosses between members of the tomato clade (Solanum section Lycopersicon, Solanaceae). Am J Bot 102: 1–10 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
