

Widespread polycistronic gene expression in green algae
- PMID: 33579822
- PMCID: PMC7896298
- DOI: 10.1073/pnas.2017714118
Widespread polycistronic gene expression in green algae
Abstract
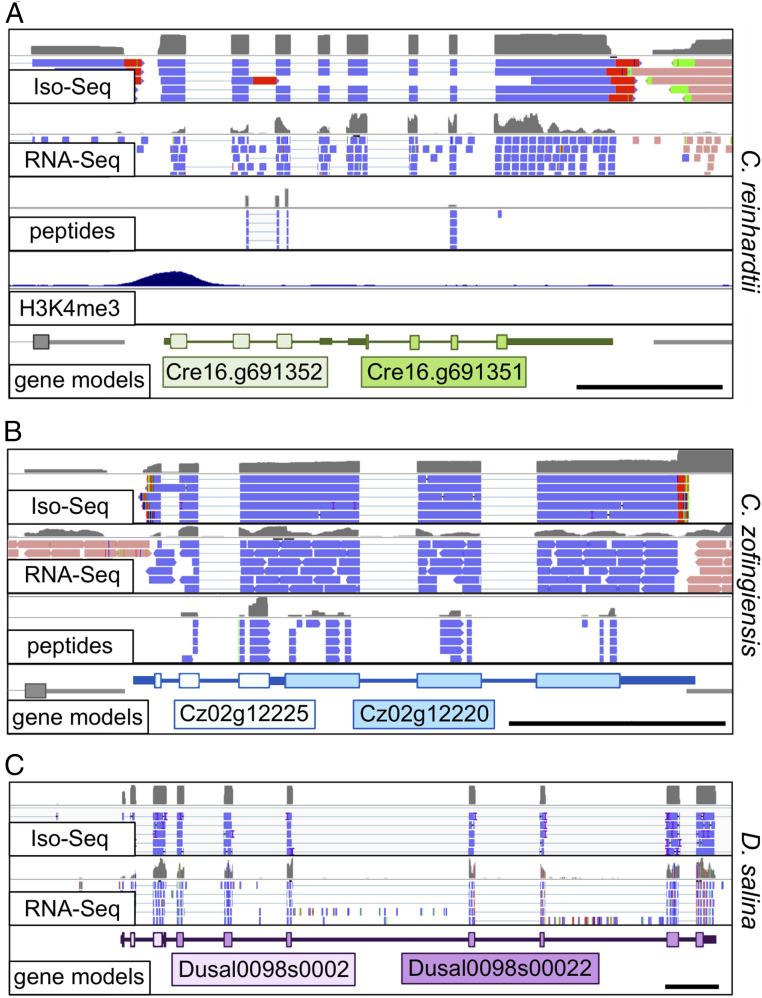
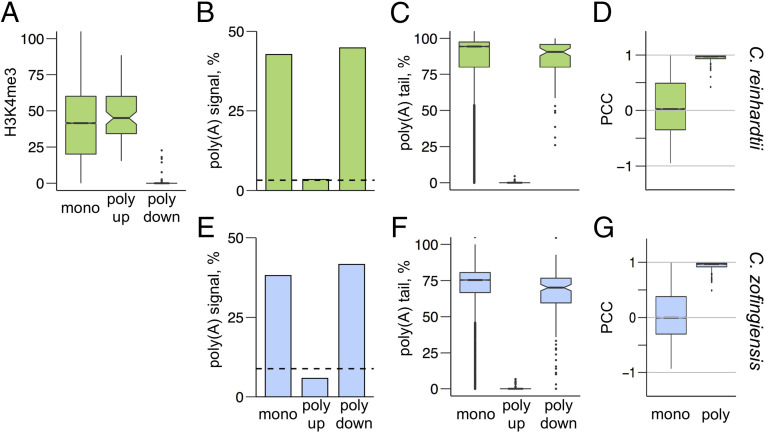
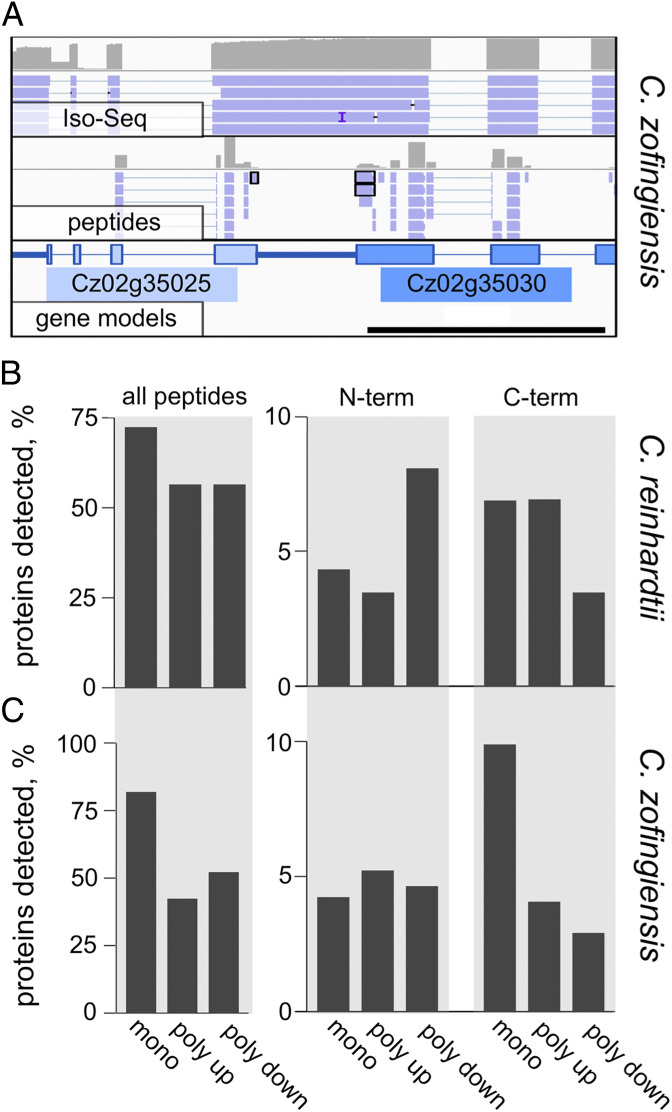
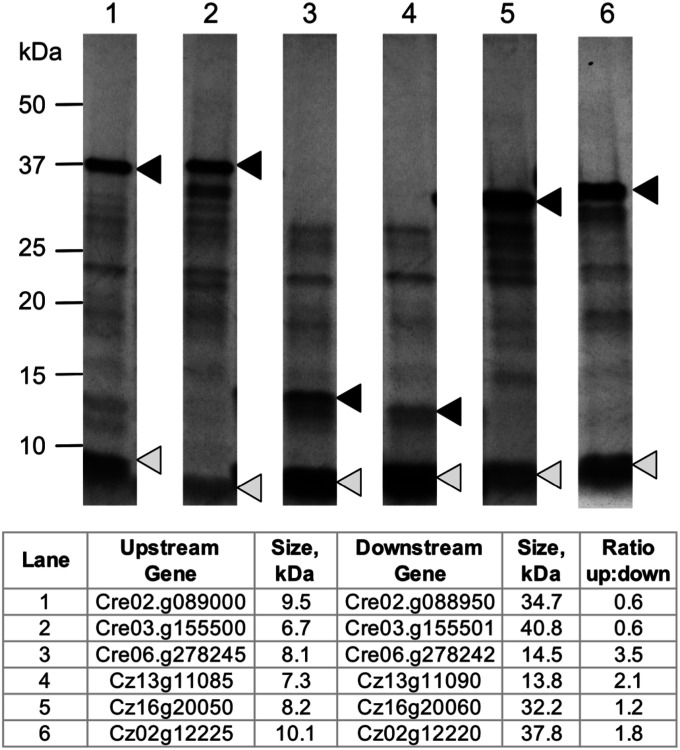
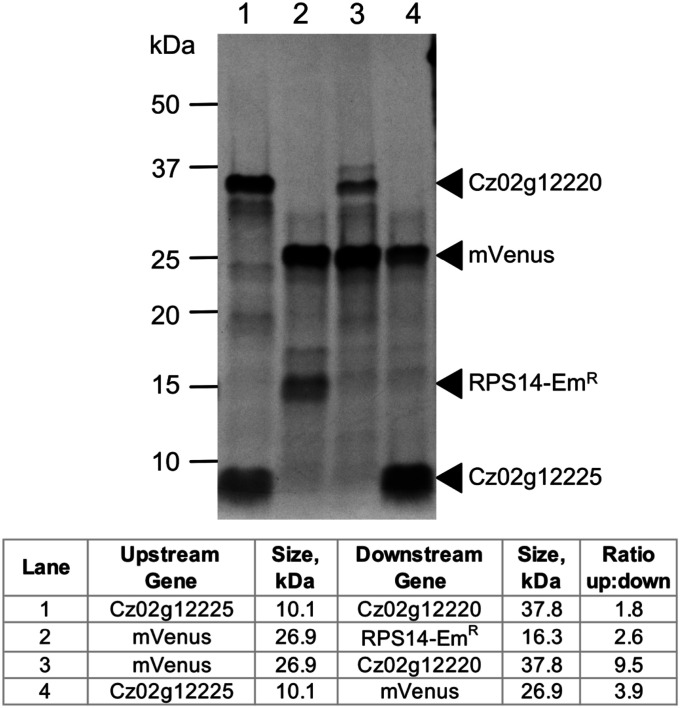
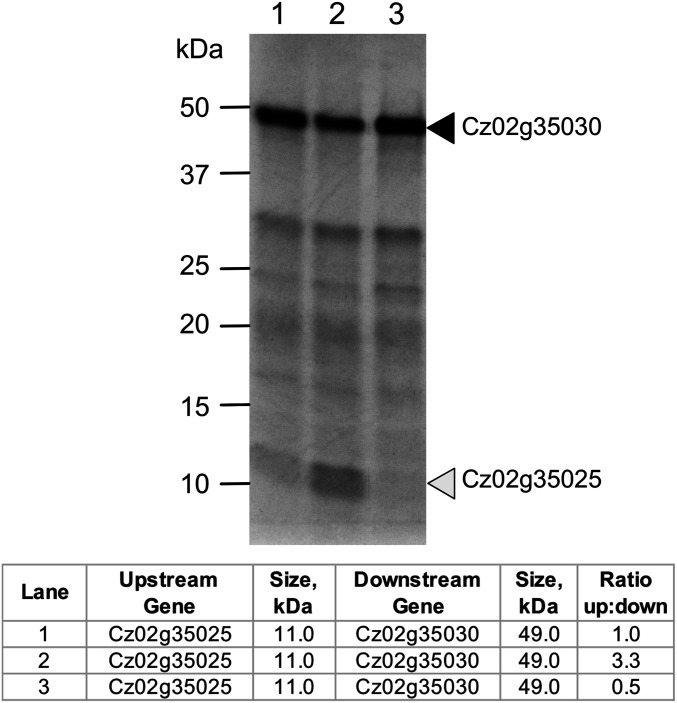
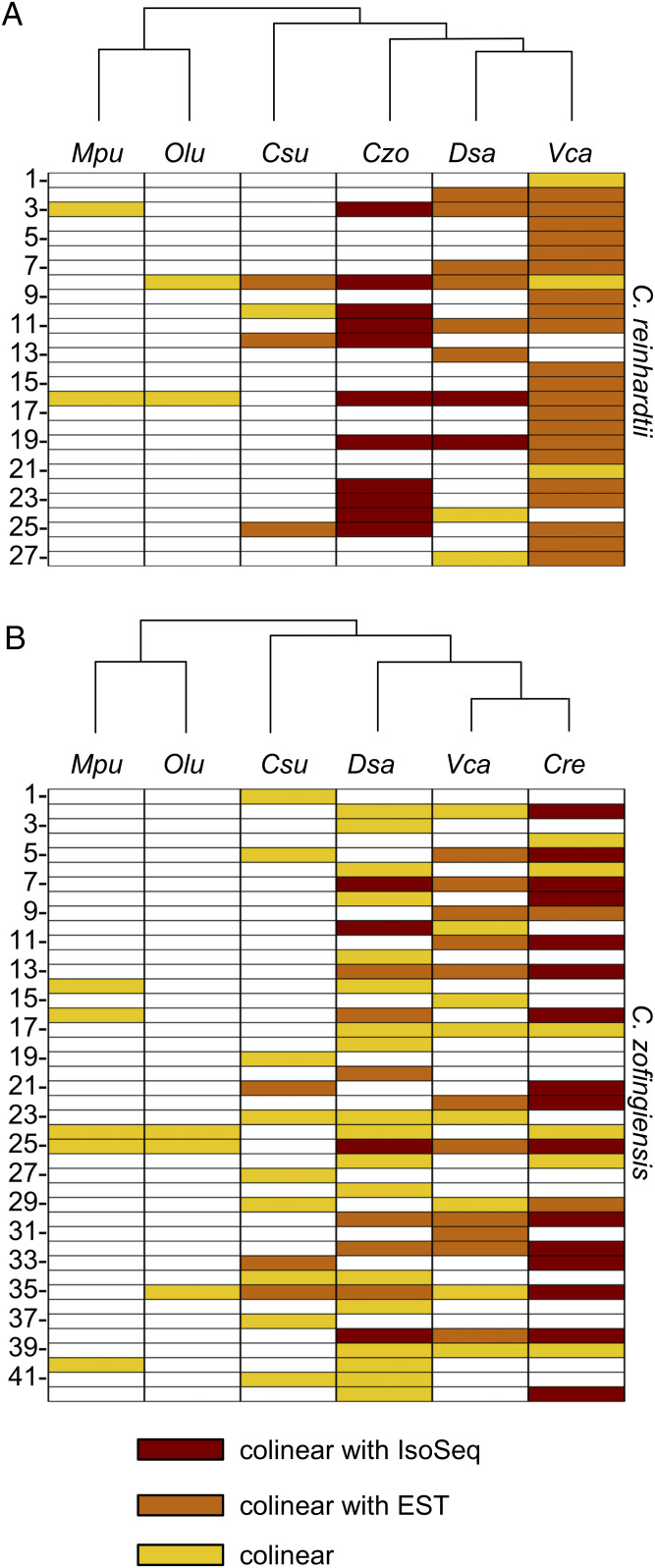
Polycistronic gene expression, common in prokaryotes, was thought to be extremely rare in eukaryotes. The development of long-read sequencing of full-length transcript isomers (Iso-Seq) has facilitated a reexamination of that dogma. Using Iso-Seq, we discovered hundreds of examples of polycistronic expression of nuclear genes in two divergent species of green algae: Chlamydomonas reinhardtii and Chromochloris zofingiensis Here, we employ a range of independent approaches to validate that multiple proteins are translated from a common transcript for hundreds of loci. A chromatin immunoprecipitation analysis using trimethylation of lysine 4 on histone H3 marks confirmed that transcription begins exclusively at the upstream gene. Quantification of polyadenylated [poly(A)] tails and poly(A) signal sequences confirmed that transcription ends exclusively after the downstream gene. Coexpression analysis found nearly perfect correlation for open reading frames (ORFs) within polycistronic loci, consistent with expression in a shared transcript. For many polycistronic loci, terminal peptides from both ORFs were identified from proteomics datasets, consistent with independent translation. Synthetic polycistronic gene pairs were transcribed and translated in vitro to recapitulate the production of two distinct proteins from a common transcript. The relative abundance of these two proteins can be modified by altering the Kozak-like sequence of the upstream gene. Replacement of the ORFs with selectable markers or reporters allows production of such heterologous proteins, speaking to utility in synthetic biology approaches. Conservation of a significant number of polycistronic gene pairs between C. reinhardtii, C. zofingiensis, and five other species suggests that this mechanism may be evolutionarily ancient and biologically important in the green algal lineage.
Keywords: bicistronic; dicistronic; leaky ribosome scanning; transcriptome; uORFs.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
Competing interest statement: S.D.G. and S.S.M. have filed a disclosure entitled “Expressing Multiple Genes from a Single Transcript in Algae and Plants.”
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
