

Coordinated changes in cellular behavior ensure the lifelong maintenance of the hippocampal stem cell population
- PMID: 33581058
- PMCID: PMC8110946
- DOI: 10.1016/j.stem.2021.01.003
Coordinated changes in cellular behavior ensure the lifelong maintenance of the hippocampal stem cell population
Abstract
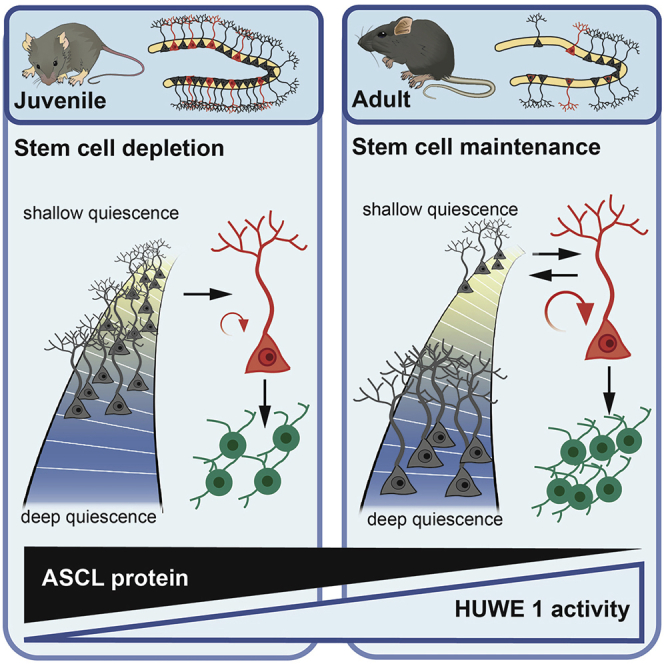
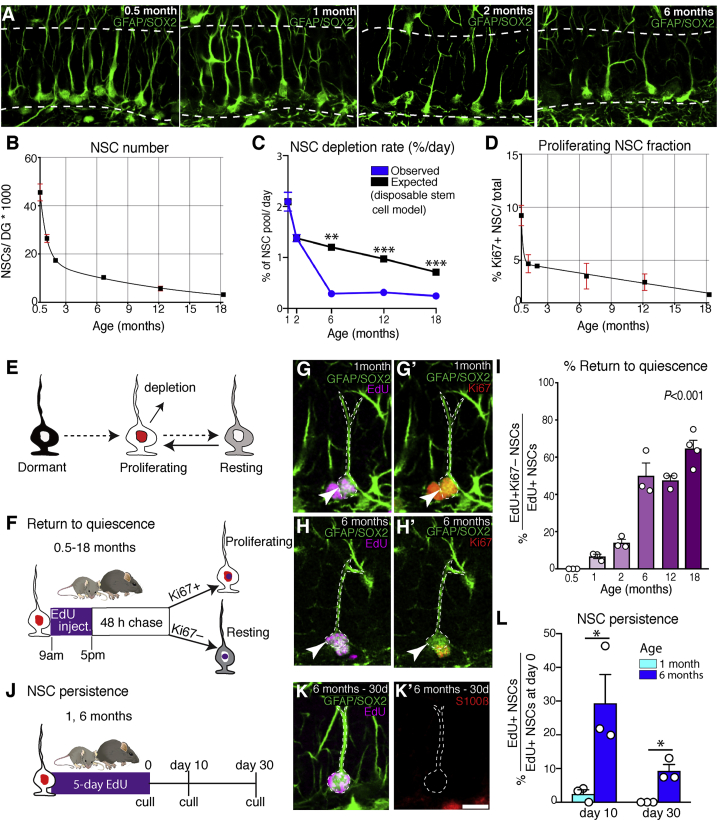
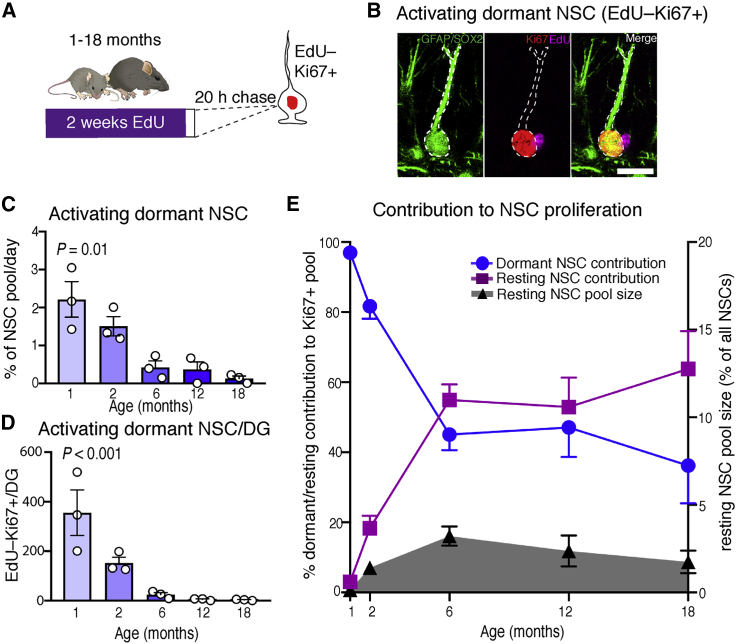
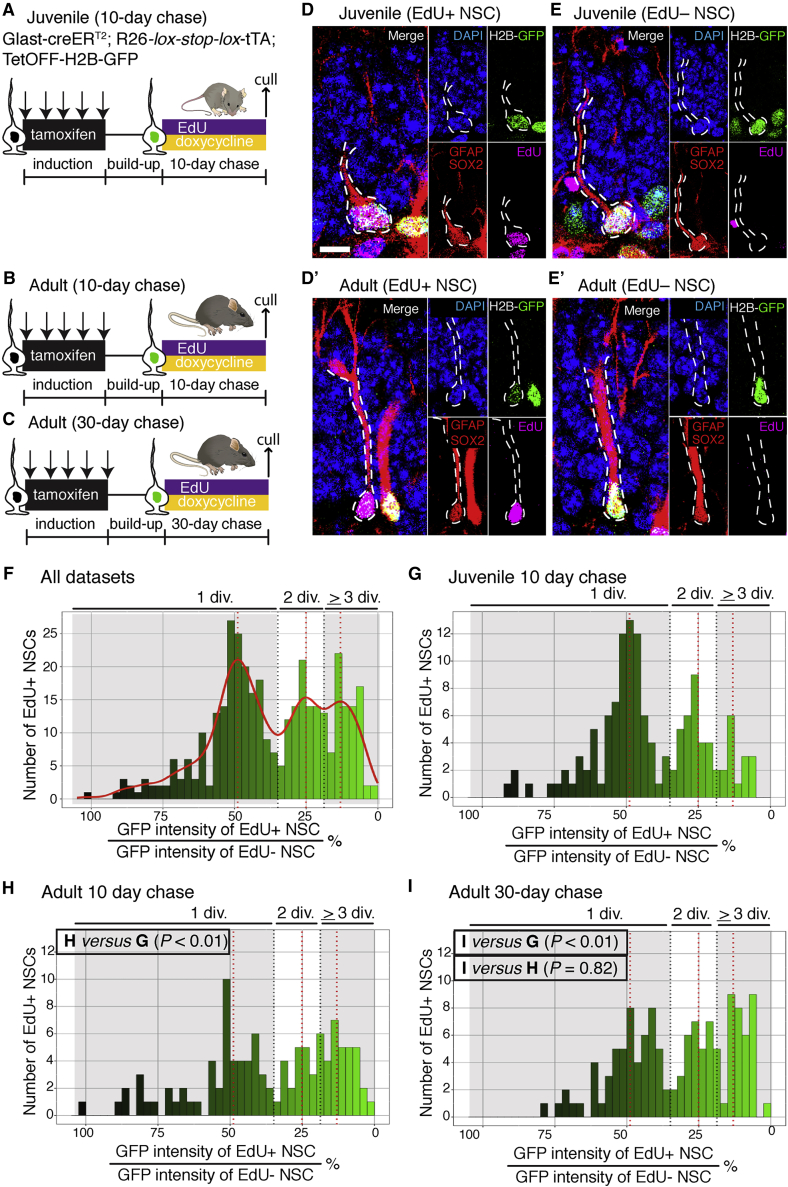
Neural stem cell numbers fall rapidly in the hippocampus of juvenile mice but stabilize during adulthood, ensuring lifelong hippocampal neurogenesis. We show that this stabilization of stem cell numbers in young adults is the result of coordinated changes in stem cell behavior. Although proliferating neural stem cells in juveniles differentiate rapidly, they increasingly return to a resting state of shallow quiescence and progress through additional self-renewing divisions in adulthood. Single-cell transcriptomics, modeling, and label retention analyses indicate that resting cells have a higher activation rate and greater contribution to neurogenesis than dormant cells, which have not left quiescence. These changes in stem cell behavior result from a progressive reduction in expression of the pro-activation protein ASCL1 because of increased post-translational degradation. These cellular mechanisms help reconcile current contradictory models of hippocampal neural stem cell (NSC) dynamics and may contribute to the different rates of decline of hippocampal neurogenesis in mammalian species, including humans.
Keywords: Ascl1; Huwe1; age; dormant; hippocampus; neural stem cell; neurogenesis; quiescence; resting.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
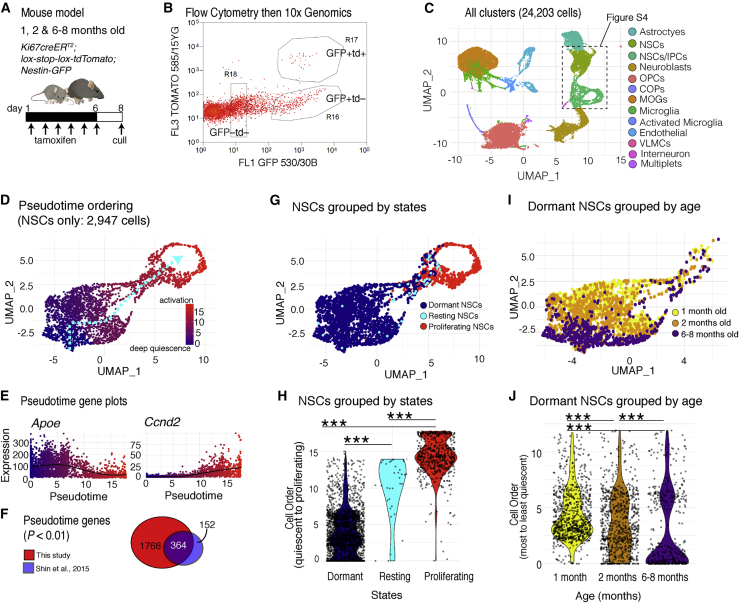
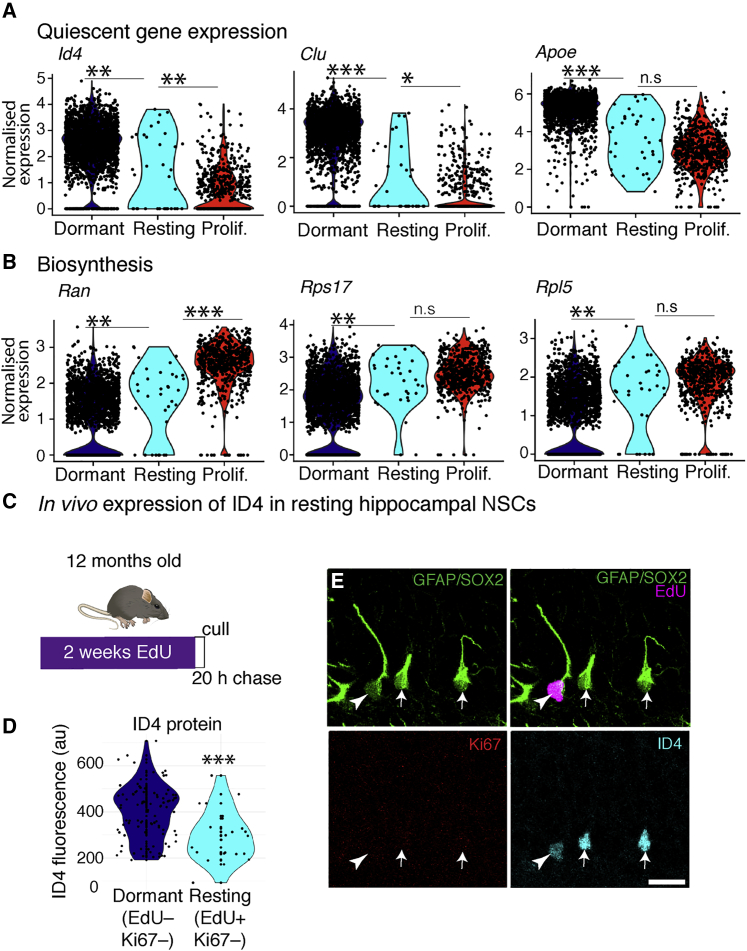
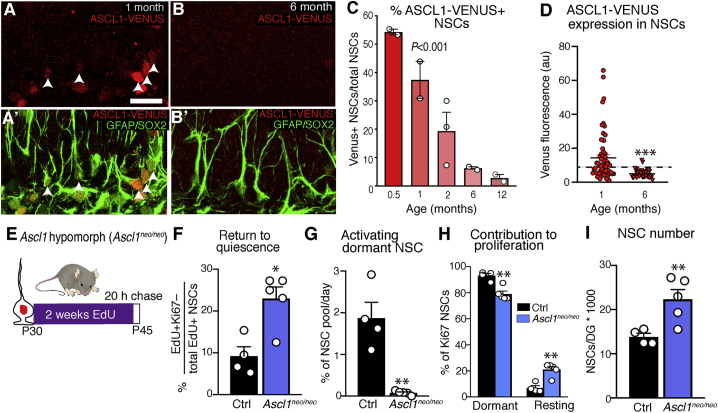
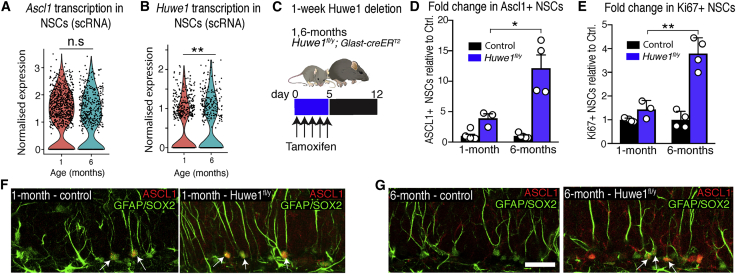
Comment in
-
The future belongs to those who prepare for it today.Cell Stem Cell. 2021 May 6;28(5):783-785. doi: 10.1016/j.stem.2021.04.014. Cell Stem Cell. 2021. PMID: 33961757
References
-
- Ali F.R., Cheng K., Kirwan P., Metcalfe S., Livesey F.J., Barker R.A., Philpott A. The phosphorylation status of Ascl1 is a key determinant of neuronal differentiation and maturation in vivo and in vitro. Development. 2014;141:2216–2224. - PubMed
-
- Ameijeiras-Alonso J., Crujeiras R.M., Rodríguez-Casal A. Multimode: An R Package for Mode Assessment. arXiv. 2018 https://arxiv.org/abs/1803.00472 arXiv:1803.00472.
-
- Barker N., Bartfeld S., Clevers H. Tissue-resident adult stem cell populations of rapidly self-renewing organs. Cell Stem Cell. 2010;7:656–670. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
