

Self-Amplifying Pestivirus Replicon RNA Encoding Influenza Virus Nucleoprotein and Hemagglutinin Promote Humoral and Cellular Immune Responses in Pigs
- PMID: 33584723
- PMCID: PMC7877248
- DOI: 10.3389/fimmu.2020.622385
Self-Amplifying Pestivirus Replicon RNA Encoding Influenza Virus Nucleoprotein and Hemagglutinin Promote Humoral and Cellular Immune Responses in Pigs
Abstract
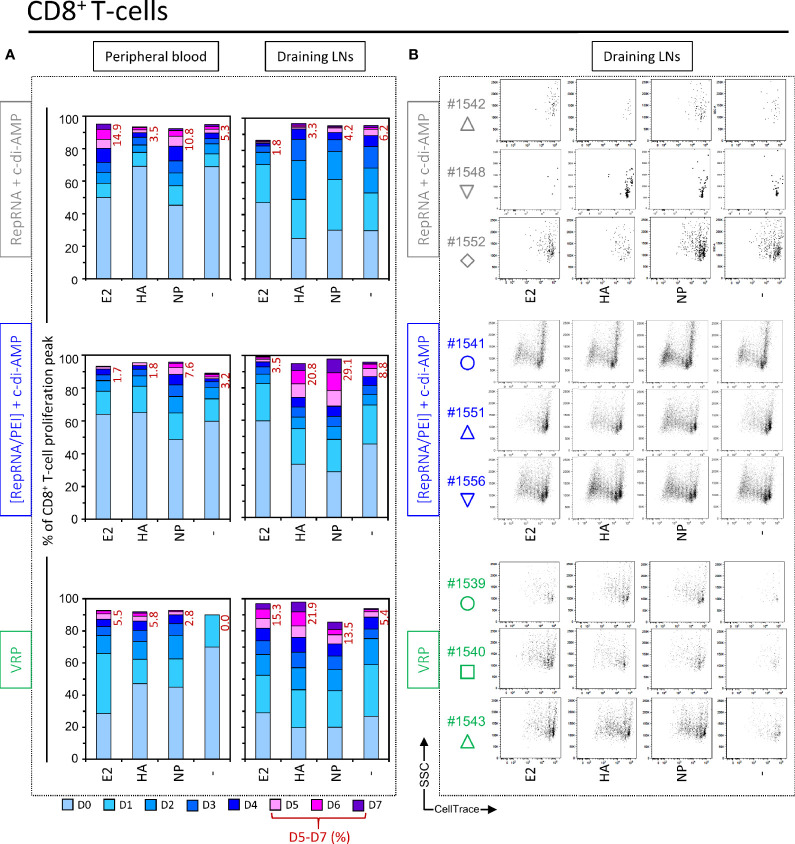
Self-amplifying replicon RNA (RepRNA) promotes expansion of mRNA templates encoding genes of interest through their replicative nature, thus providing increased antigen payloads. RepRNA derived from the non-cytopathogenic classical swine fever virus (CSFV) targets monocytes and dendritic cells (DCs), potentially promoting prolonged antigen expression in the DCs, contrasting with cytopathogenic RepRNA. We engineered pestivirus RepRNA constructs encoding influenza virus H5N1 (A/chicken/Yamaguchi/7/2004) nucleoprotein (Rep-NP) or hemagglutinin (Rep-HA). The inherent RNase-sensitivity of RepRNA had to be circumvented to ensure efficient delivery to DCs for intracellular release and RepRNA translation; we have reported how only particular synthetic delivery vehicle formulations are appropriate. The question remained concerning RepRNA packaged in virus replicon particles (VRPs); we have now compared an efficient polyethylenimine (PEI)-based formulation (polyplex) with VRP-delivery as well as naked RepRNA co-administered with the potent bis-(3',5')-cyclic dimeric adenosine monophosphate (c-di-AMP) adjuvant. All formulations contained a Rep-HA/Rep-NP mix, to assess the breadth of both humoral and cell-mediated defences against the influenza virus antigens. Assessment employed pigs for their close immunological relationship to humans, and as natural hosts for influenza virus. Animals receiving the VRPs, as well as PEI-delivered RepRNA, displayed strong humoral and cellular responses against both HA and NP, but with VRPs proving to be more efficacious. In contrast, naked RepRNA plus c-di-AMP could induce only low-level immune responses, in one out of five pigs. In conclusion, RepRNA encoding different influenza virus antigens are efficacious for inducing both humoral and cellular immune defences in pigs. Comparisons showed that packaging within VRP remains the most efficacious for delivery leading to induction of immune defences; however, this technology necessitates employment of expensive complementing cell cultures, and VRPs do not target human cells. Therefore, choosing the appropriate synthetic delivery vehicle still offers potential for rapid vaccine design, particularly in the context of the current coronavirus pandemic.
Keywords: c-di-AMP adjuvant; humoral and cellular immune response; influenza vaccines; polyplexes; self-amplifying replicon RNA; virus replicon particle.
Copyright © 2021 Démoulins, Ruggli, Gerber, Thomann-Harwood, Ebensen, Schulze, Guzmán and McCullough.
Conflict of interest statement
Application of delivery vehicles for the delivery of RepRNA vaccines to DCs (23) using replicons derived from classical swine fever virus, as employed in this paper, has been filed for patents in Europe, USA, Canada, and India, with priority date of 2008. The filing was by the authors KM and NR, together with Jon Duri Tratschin (all three as inventors) (WO 2009146867) (6), and assigned to their employer – the Institute of Virology and Immunology. This does not alter the authors’ adherence to the policies of sharing data and materials. CG and TE are named as inventors in a patent application covering the use of c-di-AMP as adjuvant (PCT/EP 2006010693). The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
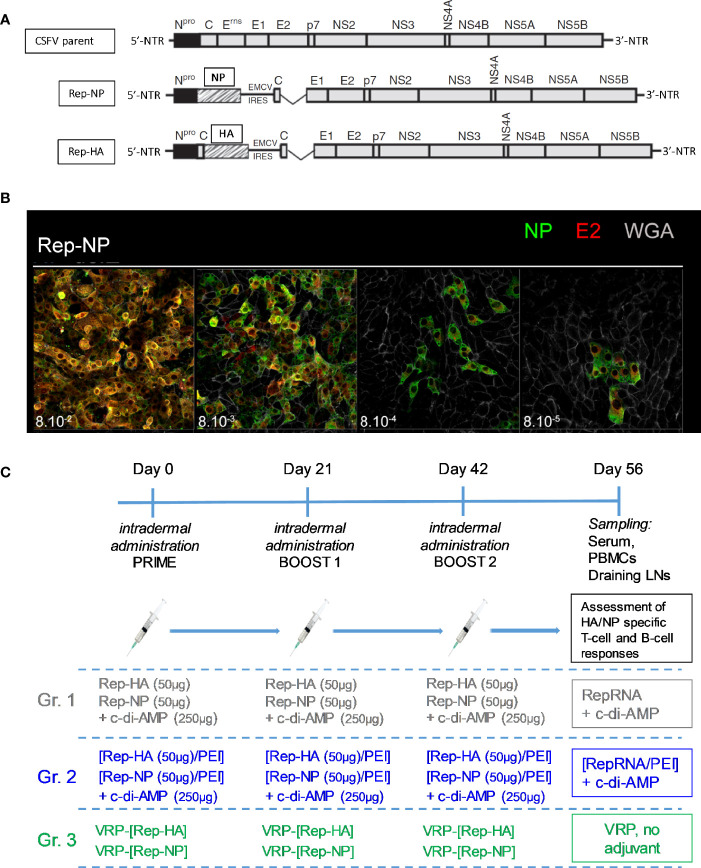
Similar articles
-
Polyethylenimine-based polyplex delivery of self-replicating RNA vaccines.Nanomedicine. 2016 Apr;12(3):711-722. doi: 10.1016/j.nano.2015.11.001. Epub 2015 Dec 1. Nanomedicine. 2016. PMID: 26592962
-
Self-replicating RNA vaccine functionality modulated by fine-tuning of polyplex delivery vehicle structure.J Control Release. 2017 Nov 28;266:256-271. doi: 10.1016/j.jconrel.2017.09.018. Epub 2017 Sep 19. J Control Release. 2017. PMID: 28935594
-
Self-Amplifying Replicon RNA Delivery to Dendritic Cells by Cationic Lipids.Mol Ther Nucleic Acids. 2018 Sep 7;12:118-134. doi: 10.1016/j.omtn.2018.04.019. Epub 2018 May 4. Mol Ther Nucleic Acids. 2018. PMID: 30195751 Free PMC article.
-
Self-Replicating RNA Vaccine Delivery to Dendritic Cells.Methods Mol Biol. 2017;1499:37-75. doi: 10.1007/978-1-4939-6481-9_3. Methods Mol Biol. 2017. PMID: 27987142 Review.
-
Progress on adenovirus-vectored universal influenza vaccines.Hum Vaccin Immunother. 2015;11(5):1209-22. doi: 10.1080/21645515.2015.1016674. Hum Vaccin Immunother. 2015. PMID: 25876176 Free PMC article. Review.
Cited by
-
The Promise and Challenges of Cyclic Dinucleotides as Molecular Adjuvants for Vaccine Development.Vaccines (Basel). 2021 Aug 17;9(8):917. doi: 10.3390/vaccines9080917. Vaccines (Basel). 2021. PMID: 34452042 Free PMC article. Review.
-
Construction and evaluation of a self-replicative RNA vaccine against SARS-CoV-2 using yellow fever virus replicon.PLoS One. 2022 Oct 20;17(10):e0274829. doi: 10.1371/journal.pone.0274829. eCollection 2022. PLoS One. 2022. PMID: 36264936 Free PMC article.
-
Packaging, Purification, and Titration of Replication-Deficient Semliki Forest Virus-Derived Particles as a Self-Amplifying mRNA Vaccine Vector.Iran Biomed J. 2022 Jul 1;26(4):269-78. doi: 10.52547/ibj.3535. Iran Biomed J. 2022. PMID: 35468712 Free PMC article.
-
Assessment of Immune Responses to a Trivalent Pichinde Virus-Vectored Vaccine Expressing Hemagglutinin Genes from Three Co-Circulating Influenza A Virus Subtypes in Pigs.Vaccines (Basel). 2023 Dec 2;11(12):1806. doi: 10.3390/vaccines11121806. Vaccines (Basel). 2023. PMID: 38140210 Free PMC article.
-
New Generation Self-Replicating RNA Vaccines Derived from Pestivirus Genome.Methods Mol Biol. 2024;2786:89-133. doi: 10.1007/978-1-0716-3770-8_4. Methods Mol Biol. 2024. PMID: 38814391
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous