

Haspin Modulates the G2/M Transition Delay in Response to Polarization Failures in Budding Yeast
- PMID: 33585466
- PMCID: PMC7876276
- DOI: 10.3389/fcell.2020.625717
Haspin Modulates the G2/M Transition Delay in Response to Polarization Failures in Budding Yeast
Abstract
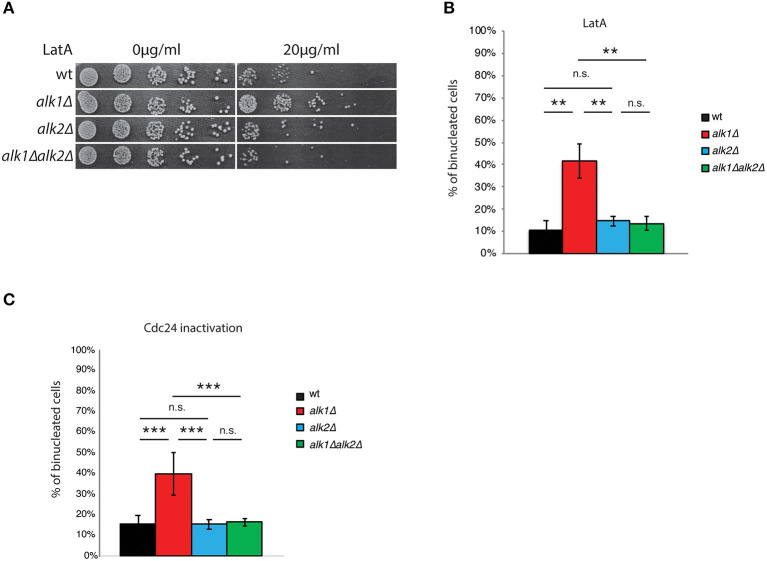
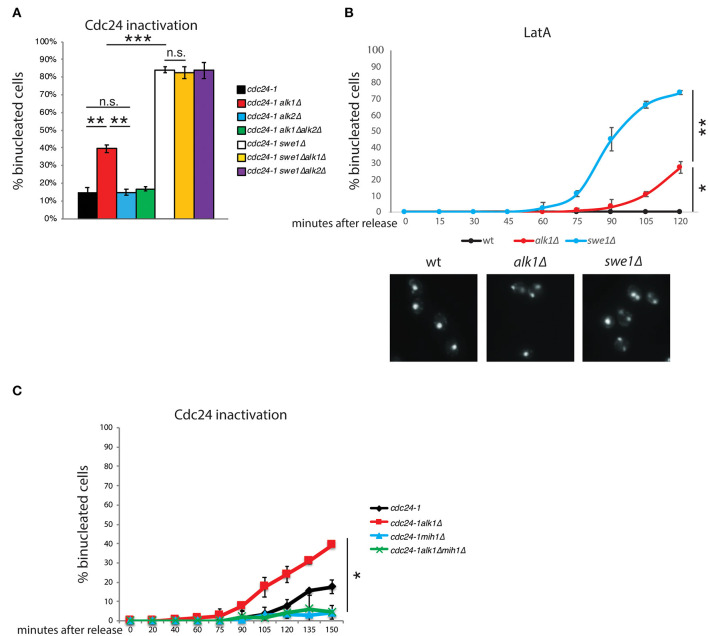
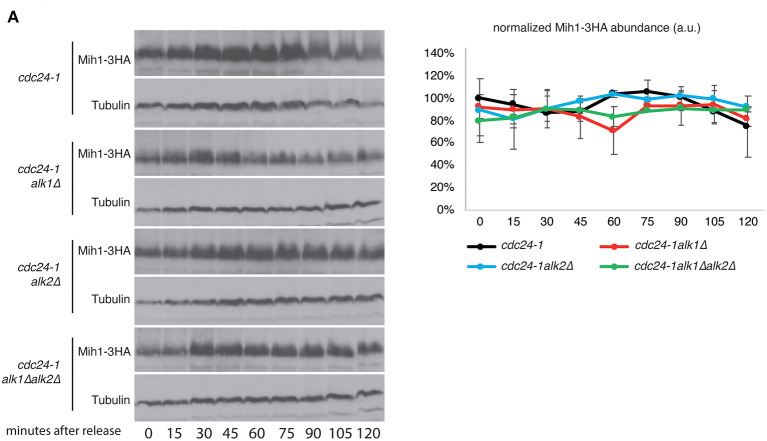
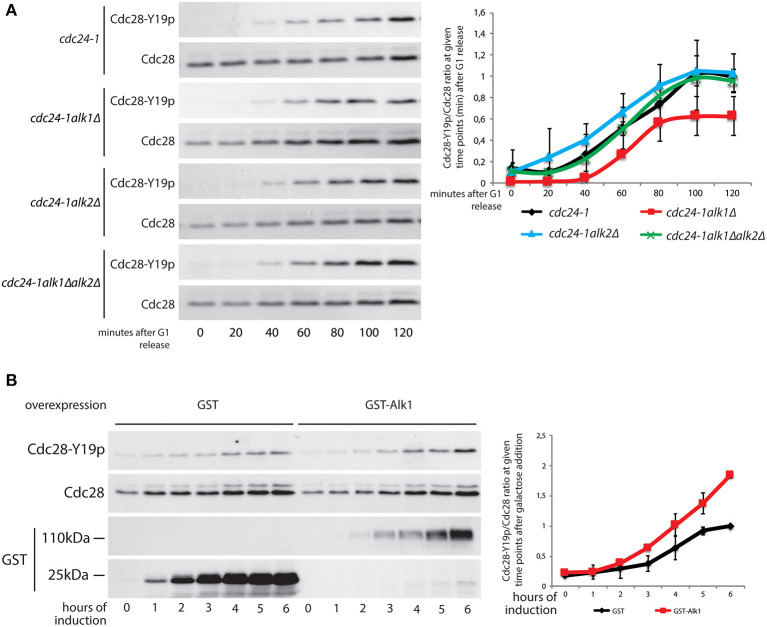
Symmetry breaking by cellular polarization is an exquisite requirement for the cell-cycle of Saccharomyces cerevisiae cells, as it allows bud emergence and growth. This process is based on the formation of polarity clusters at the incipient bud site, first, and the bud tip later in the cell-cycle, that overall promote bud emission and growth. Given the extreme relevance of this process, a surveillance mechanism, known as the morphogenesis checkpoint, has evolved to coordinate the formation of the bud and cell cycle progression, delaying mitosis in the presence of morphogenetic problems. The atypical protein kinase haspin is responsible for histone H3-T3 phosphorylation and, in yeast, for resolution of polarity clusters in mitosis. Here, we report a novel role for haspin in the regulation of the morphogenesis checkpoint in response to polarity insults. Particularly, we show that cells lacking the haspin ortholog Alk1 fail to achieve sustained checkpoint activation and enter mitosis even in the absence of a bud. In alk1Δ cells, we report a reduced phosphorylation of Cdc28-Y19, which stems from a premature activation of the Mih1 phosphatase. Overall, the data presented in this work define yeast haspin as a novel regulator of the morphogenesis checkpoint in Saccharomyces cerevisiae, where it monitors polarity establishment and it couples bud emergence to the G2/M cell cycle transition.
Keywords: Saccharomyces cerevisiae; actin cytoskeleton; cell cycle; mitosis; morphogenesis checkpoint; polarization.
Copyright © 2021 Galli, Diani, Quadri, Nespoli, Galati, Panigada, Plevani and Muzi-Falconi.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Conservation of mechanisms controlling entry into mitosis: budding yeast wee1 delays entry into mitosis and is required for cell size control.Curr Biol. 2003 Feb 18;13(4):264-75. doi: 10.1016/s0960-9822(03)00049-6. Curr Biol. 2003. PMID: 12593792
-
Yeast haspin kinase regulates polarity cues necessary for mitotic spindle positioning and is required to tolerate mitotic arrest.Dev Cell. 2013 Sep 16;26(5):483-95. doi: 10.1016/j.devcel.2013.07.013. Epub 2013 Aug 22. Dev Cell. 2013. PMID: 23973165
-
Cdc28 tyrosine phosphorylation and the morphogenesis checkpoint in budding yeast.Mol Biol Cell. 1996 Nov;7(11):1657-66. doi: 10.1091/mbc.7.11.1657. Mol Biol Cell. 1996. PMID: 8930890 Free PMC article.
-
Cell cycle control of morphogenesis in budding yeast.Curr Opin Genet Dev. 1995 Feb;5(1):17-23. doi: 10.1016/s0959-437x(95)90048-9. Curr Opin Genet Dev. 1995. PMID: 7749320 Review.
-
Eavesdropping on the cytoskeleton: progress and controversy in the yeast morphogenesis checkpoint.Curr Opin Microbiol. 2006 Dec;9(6):540-6. doi: 10.1016/j.mib.2006.10.004. Epub 2006 Oct 19. Curr Opin Microbiol. 2006. PMID: 17055334 Review.
Cited by
-
Phosphorylation of H3-Thr3 by Haspin Is Required for Primary Cilia Regulation.Int J Mol Sci. 2021 Jul 20;22(14):7753. doi: 10.3390/ijms22147753. Int J Mol Sci. 2021. PMID: 34299370 Free PMC article.
-
Distinct Aurora B pools at the inner centromere and kinetochore have different contributions to meiotic and mitotic chromosome segregation.Mol Biol Cell. 2023 May 1;34(5):ar43. doi: 10.1091/mbc.E23-01-0014. Epub 2023 Mar 15. Mol Biol Cell. 2023. PMID: 36920098 Free PMC article.
-
Distinct Aurora B pools at the inner centromere and kinetochore have different contributions to meiotic and mitotic chromosome segregation.bioRxiv [Preprint]. 2023 Feb 5:2023.02.05.527197. doi: 10.1101/2023.02.05.527197. bioRxiv. 2023. Update in: Mol Biol Cell. 2023 May 1;34(5):ar43. doi: 10.1091/mbc.E23-01-0014. PMID: 36778459 Free PMC article. Updated. Preprint.
-
A Haspin-ARHGAP11A axis regulates epithelial morphogenesis through Rho-ROCK dependent modulation of LIMK1-Cofilin.iScience. 2023 Sep 22;26(10):108011. doi: 10.1016/j.isci.2023.108011. eCollection 2023 Oct 20. iScience. 2023. PMID: 37841592 Free PMC article.
References
-
- Ayscough K. R., Stryker J., Pokala N., Sanders M., Crews P., Drubin D. G. (1997). High rates of actin filament turnover in budding yeast roles for actin in establishment maintenance of cell polarity revealed using the actin inhibitor latrunculin-A. J. Cell Biol. 137, 399–416. 10.1083/jcb.137.2.399 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases