

Cardiomyocyte Na+/H+ Exchanger-1 Activity Is Reduced in Hypoxia
- PMID: 33585583
- PMCID: PMC7873356
- DOI: 10.3389/fcvm.2020.617038
Cardiomyocyte Na+/H+ Exchanger-1 Activity Is Reduced in Hypoxia
Abstract
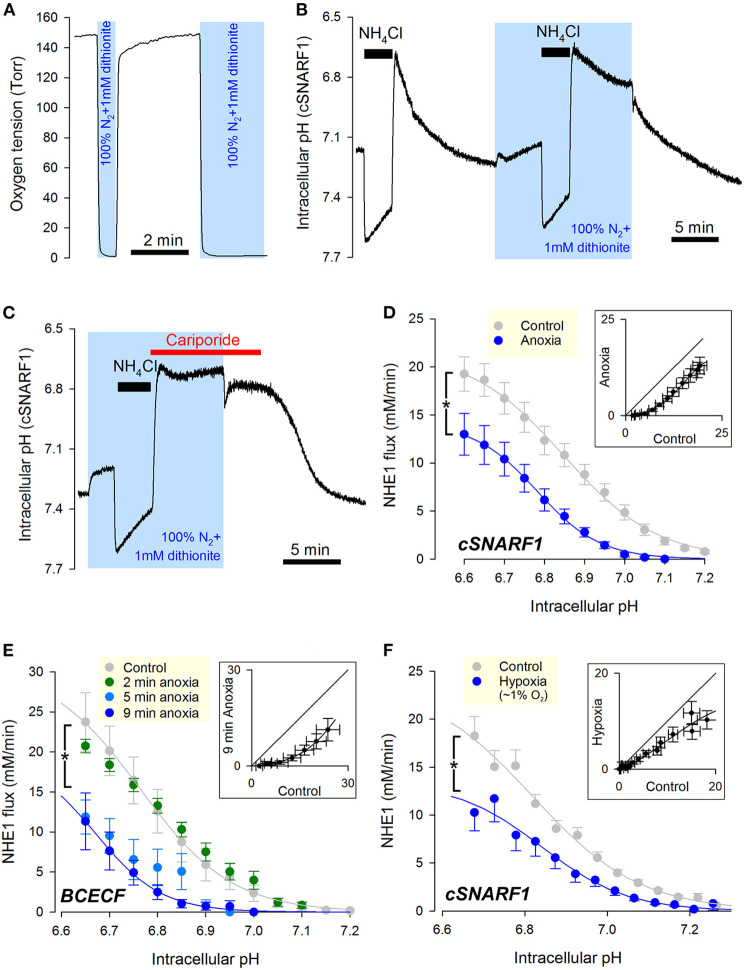
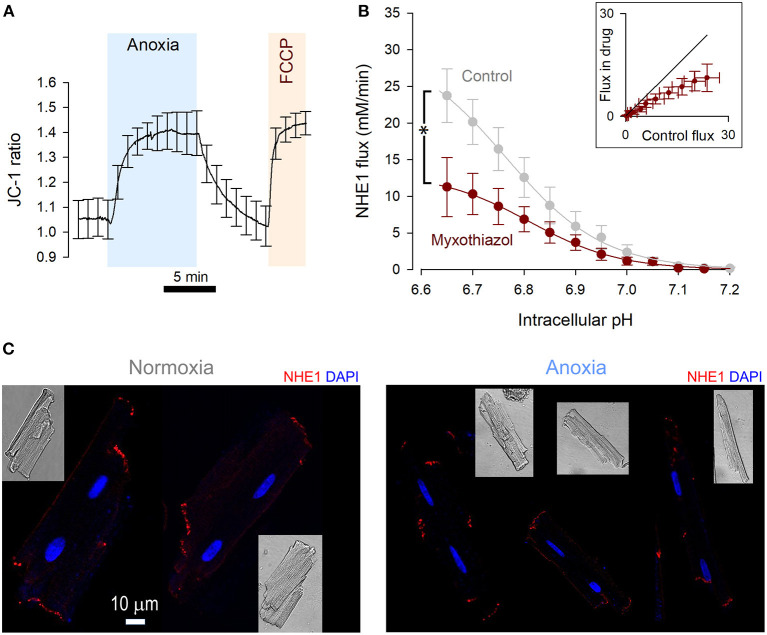
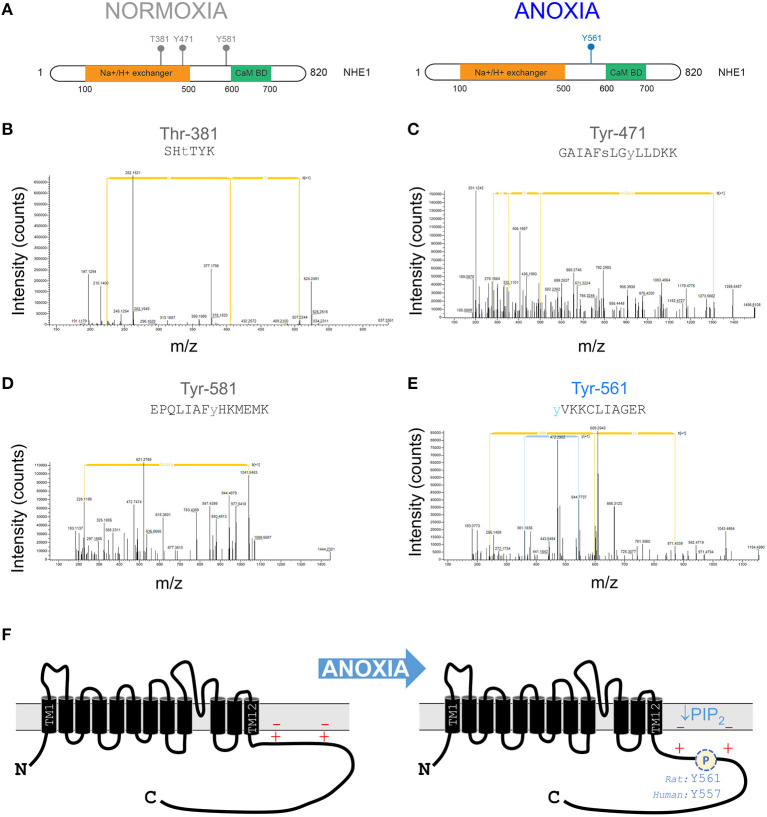
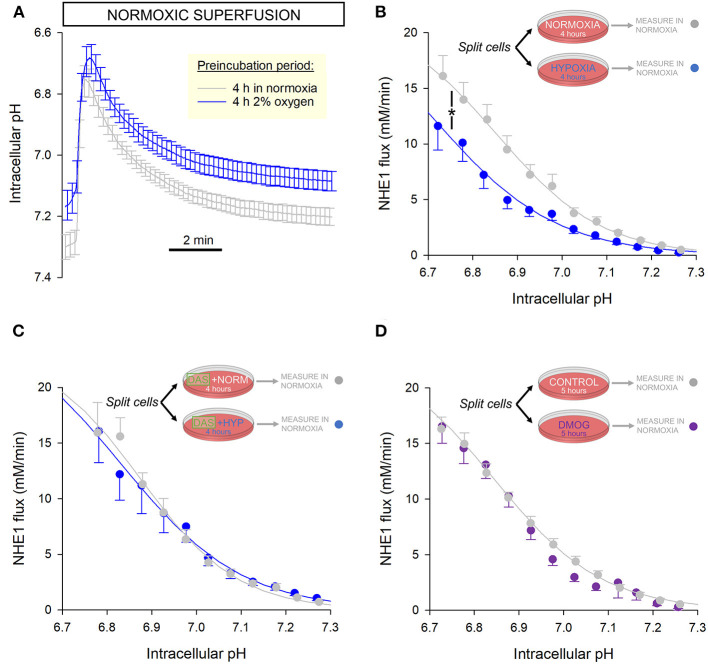
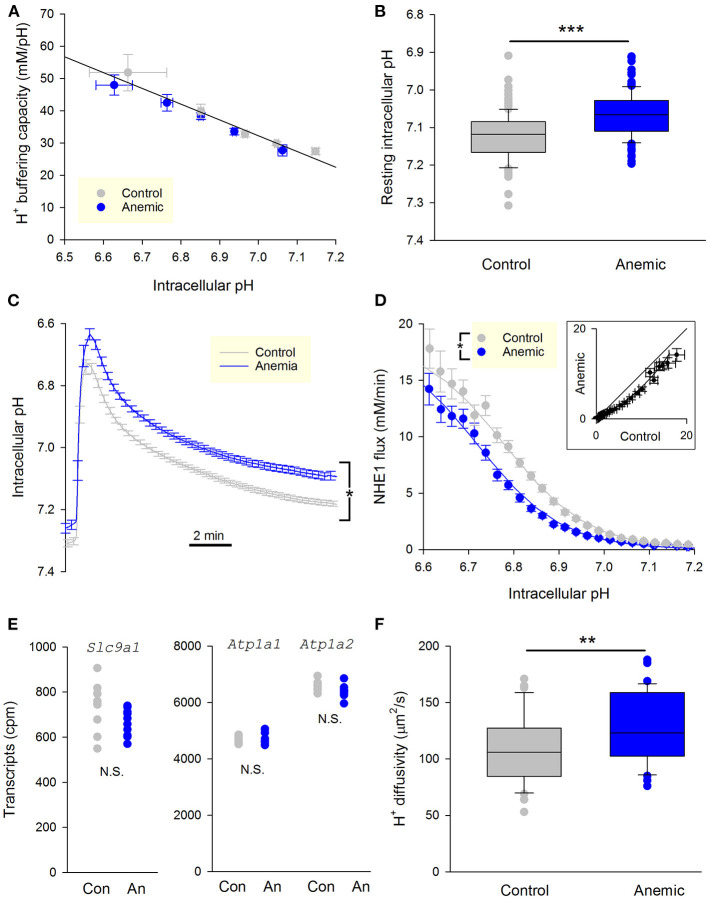
Fully-activated Na+/H+ exchanger-1 (NHE1) generates the cardiomyocyte's largest trans-membrane extrusion of H+ ions for an equimolar influx of Na+ ions. This has the desirable effect of clearing excess intracellular acidity, but comes at a large energetic premium because the exchanged Na+ ions must ultimately be extruded by the sodium pump, a process that consumes the majority of the heart's non-contractile ATP. We hypothesize that the state of NHE1 activation depends on metabolic resources, which become limiting in periods of myocardial hypoxia. To test this functionally, NHE1 activity was measured in response to in vitro and in vivo hypoxic treatments. NHE1 flux was interrogated as a function of intracellular pH by fluorescence imaging of rodent ventricular myocytes loaded with pH-sensitive dyes BCECF or cSNARF1. Anoxic superfusates promptly inhibited NHE1, tracking the time-course of mitochondrial depolarization. Mass spectrometry of NHE1 immuno-precipitated from Langendorff-perfused anoxic hearts identified Tyr-581 dephosphorylation and Tyr-561 phosphorylation. The latter residue is part of the domain that interacts with phosphatidylinositol 4,5-bisphosphate (PIP2), a membrane lipid that becomes depleted under metabolic inhibition. Tyr-561 phosphorylation is expected to electrostatically weaken this activatory interaction. To test if a period of hypoxia produces a persistent inhibition of NHE1, measurements under normoxia were performed on myocytes that had been incubated in 2% O2 for 4 h. NHE1 activity remained inhibited, but the effect was ablated in the presence of Dasatinib, an inhibitor of Abl/Src-family tyrosine kinases. Chronic tissue hypoxia in vivo, attained in a mouse model of anemic hypoxia, also resulted in persistently slower NHE1. In summary, we show that NHE1 responds to oxygen, a physiologically-relevant metabolic regulator, ostensibly to divert ATP for contraction. We describe a novel mechanism of NHE1 inhibition that may be relevant in cardiac disorders featuring altered oxygen metabolism, such as myocardial ischemia and reperfusion injury.
Keywords: ATP; NHE1; Y557; anemia; metabolism; oxygen; pH regulation; ventricle.
Copyright © 2021 Kandilci, Richards, Fournier, Şimşek, Chung, Lakhal-Littleton and Swietach.
Conflict of interest statement
YC has received personal support from Vifor Pharma for work on iron biology not related to the current study. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous