

Gain-of-function GABRB3 variants identified in vigabatrin-hypersensitive epileptic encephalopathies
- PMID: 33585817
- PMCID: PMC7869430
- DOI: 10.1093/braincomms/fcaa162
Gain-of-function GABRB3 variants identified in vigabatrin-hypersensitive epileptic encephalopathies
Abstract
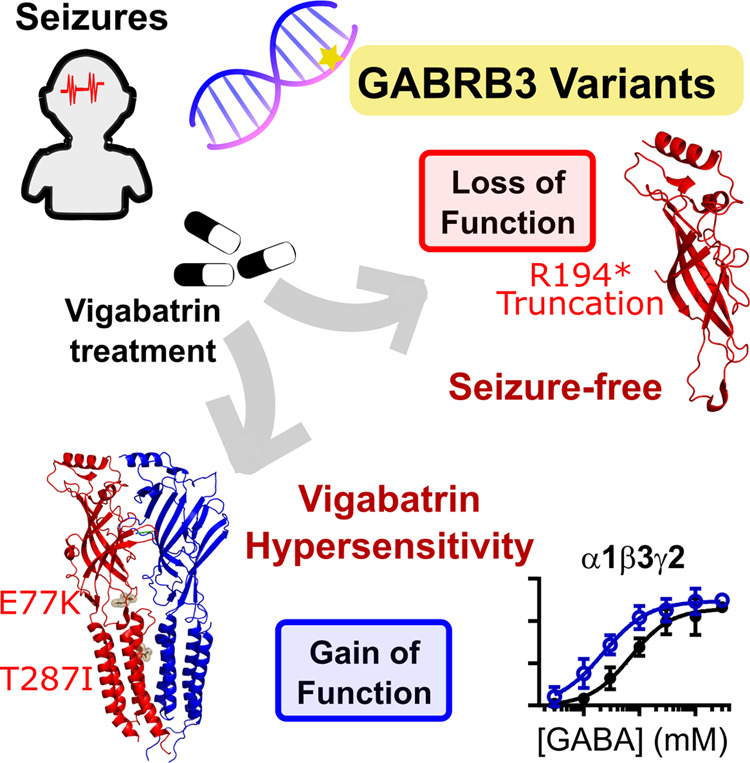
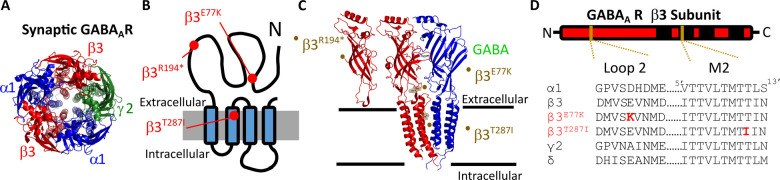
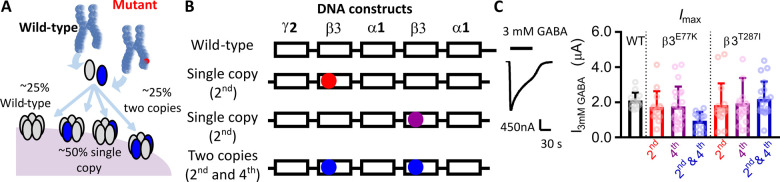
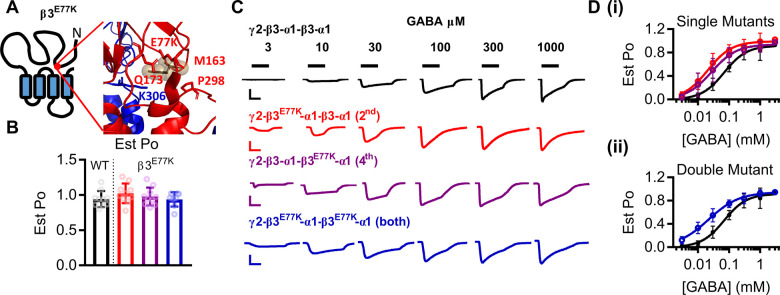
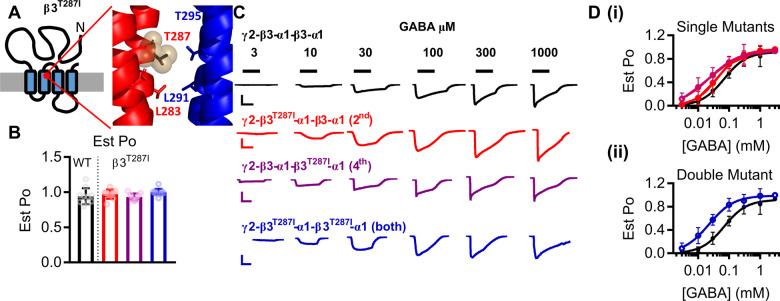
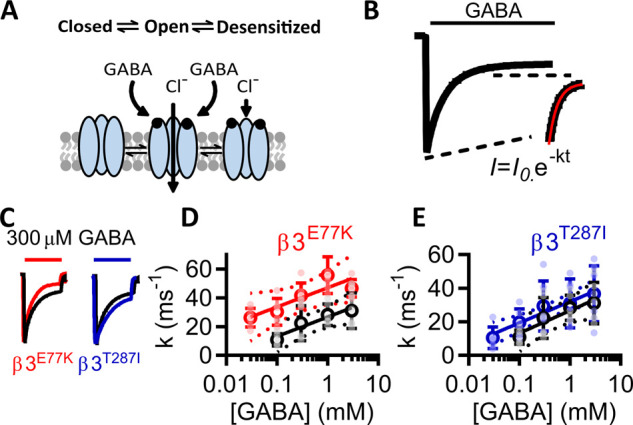
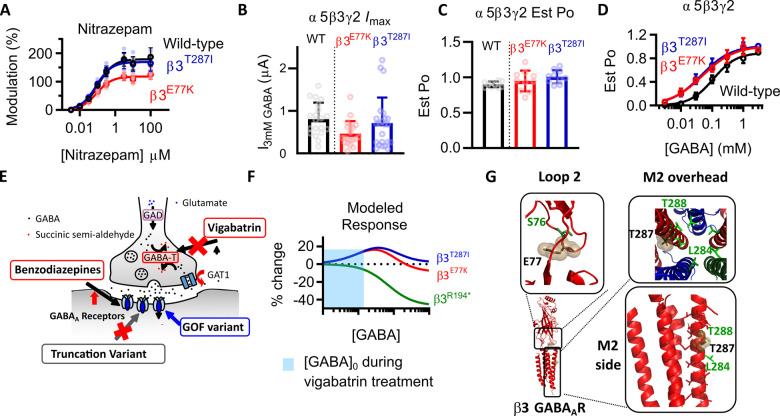
Variants in the GABRB3 gene encoding the β3-subunit of the γ-aminobutyric acid type A ( receptor are associated with various developmental and epileptic encephalopathies. Typically, these variants cause a loss-of-function molecular phenotype whereby γ-aminobutyric acid has reduced inhibitory effectiveness leading to seizures. Drugs that potentiate inhibitory GABAergic activity, such as nitrazepam, phenobarbital or vigabatrin, are expected to compensate for this and thereby reduce seizure frequency. However, vigabatrin, a drug that inhibits γ-aminobutyric acid transaminase to increase tonic γ-aminobutyric acid currents, has mixed success in treating seizures in patients with GABRB3 variants: some patients experience seizure cessation, but there is hypersensitivity in some patients associated with hypotonia, sedation and respiratory suppression. A GABRB3 variant that responds well to vigabatrin involves a truncation variant (p.Arg194*) resulting in a clear loss-of-function. We hypothesized that patients with a hypersensitive response to vigabatrin may exhibit a different γ-aminobutyric acid A receptor phenotype. To test this hypothesis, we evaluated the phenotype of de novo variants in GABRB3 (p.Glu77Lys and p.Thr287Ile) associated with patients who are clinically hypersensitive to vigabatrin. We introduced the GABRB3 p.Glu77Lys and p.Thr287Ile variants into a concatenated synaptic and extrasynaptic γ-aminobutyric acid A receptor construct, to resemble the γ-aminobutyric acid A receptor expression by a patient heterozygous for the GABRB3 variant. The mRNA of these constructs was injected into Xenopus oocytes and activation properties of each receptor measured by two-electrode voltage clamp electrophysiology. Results showed an atypical gain-of-function molecular phenotype in the GABRB3 p.Glu77Lys and p.Thr287Ile variants characterized by increased potency of γ-aminobutyric acid A without change to the estimated maximum open channel probability, deactivation kinetics or absolute currents. Modelling of the activation properties of the receptors indicated that either variant caused increased chloride flux in response to low concentrations of γ-aminobutyric acid that mediate tonic currents. We therefore propose that the hypersensitivity reaction to vigabatrin is a result of GABRB3 variants that exacerbate GABAergic tonic currents and caution is required when prescribing vigabatrin. In contrast, drug strategies increasing tonic currents in loss-of-function variants are likely to be a safe and effective therapy. This study demonstrates that functional genomics can explain beneficial and adverse anti-epileptic drug effects, and propose that vigabatrin should be considered in patients with clear loss-of-function GABRB3 variants.
Keywords: GABAA receptor; benzodiazepines; developmental and epileptic encephalopathies; gain-of-function variants; vigabatrin.
© The Author(s) (2020). Published by Oxford University Press on behalf of the Guarantors of Brain.
Figures
References
-
- Chiron C, Dulac O. The pharmacologic treatment of Dravet syndrome. Epilepsia 2011; 52: 72–5. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources