

Tet2 Inactivation Enhances the Antitumor Activity of Tumor-Infiltrating Lymphocytes
- PMID: 33589517
- PMCID: PMC8137576
- DOI: 10.1158/0008-5472.CAN-20-3213
Tet2 Inactivation Enhances the Antitumor Activity of Tumor-Infiltrating Lymphocytes
Abstract
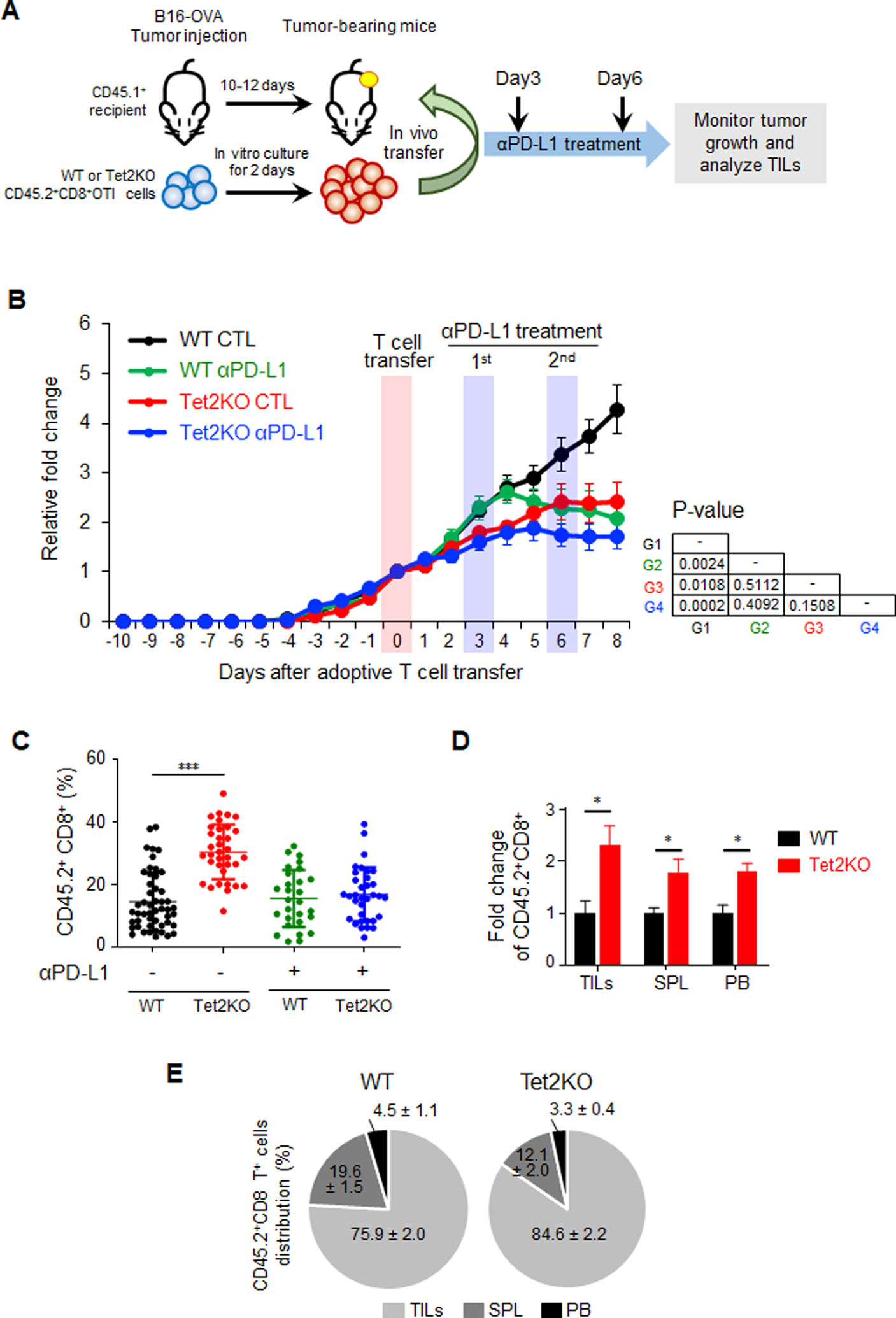
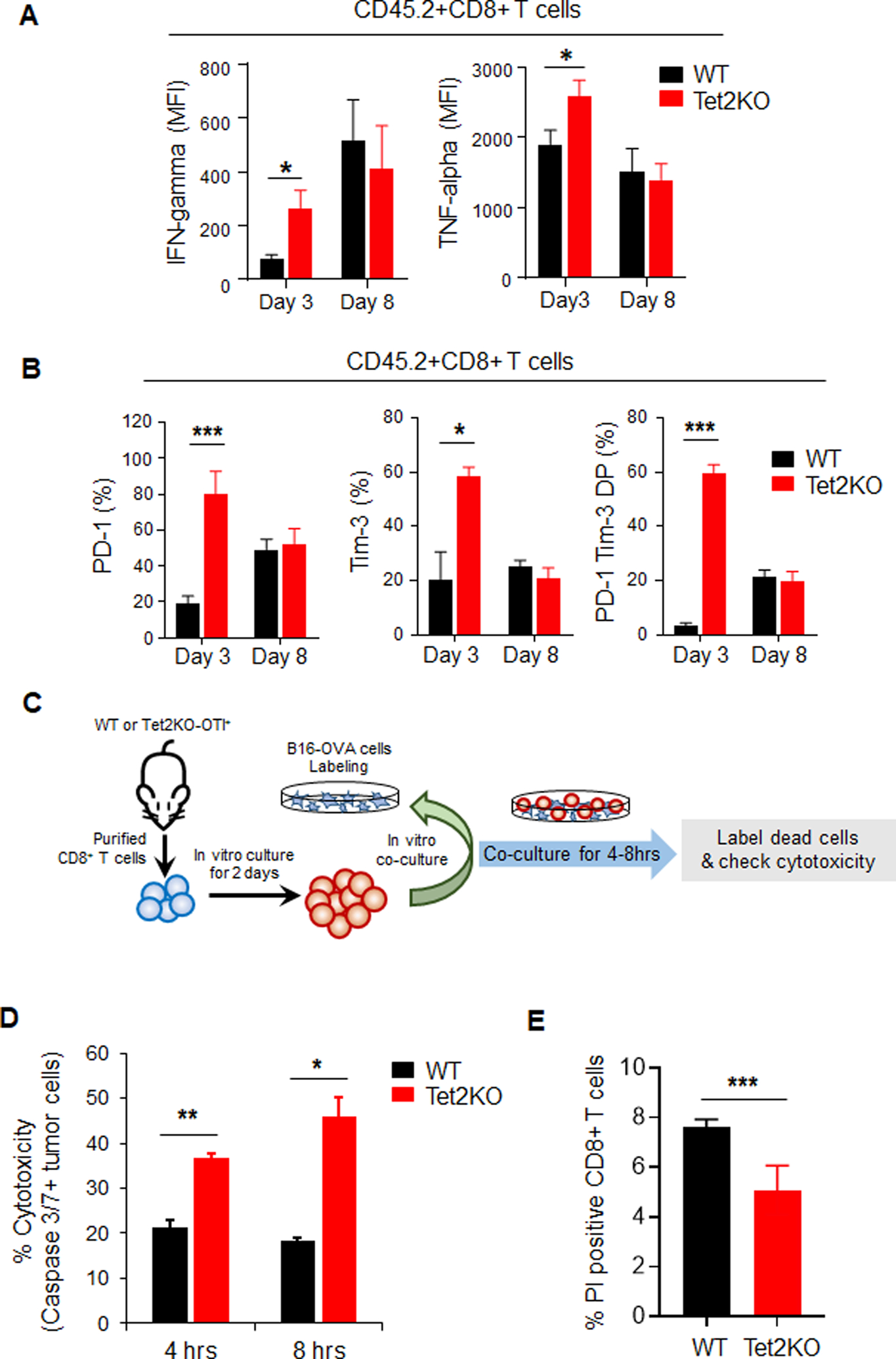
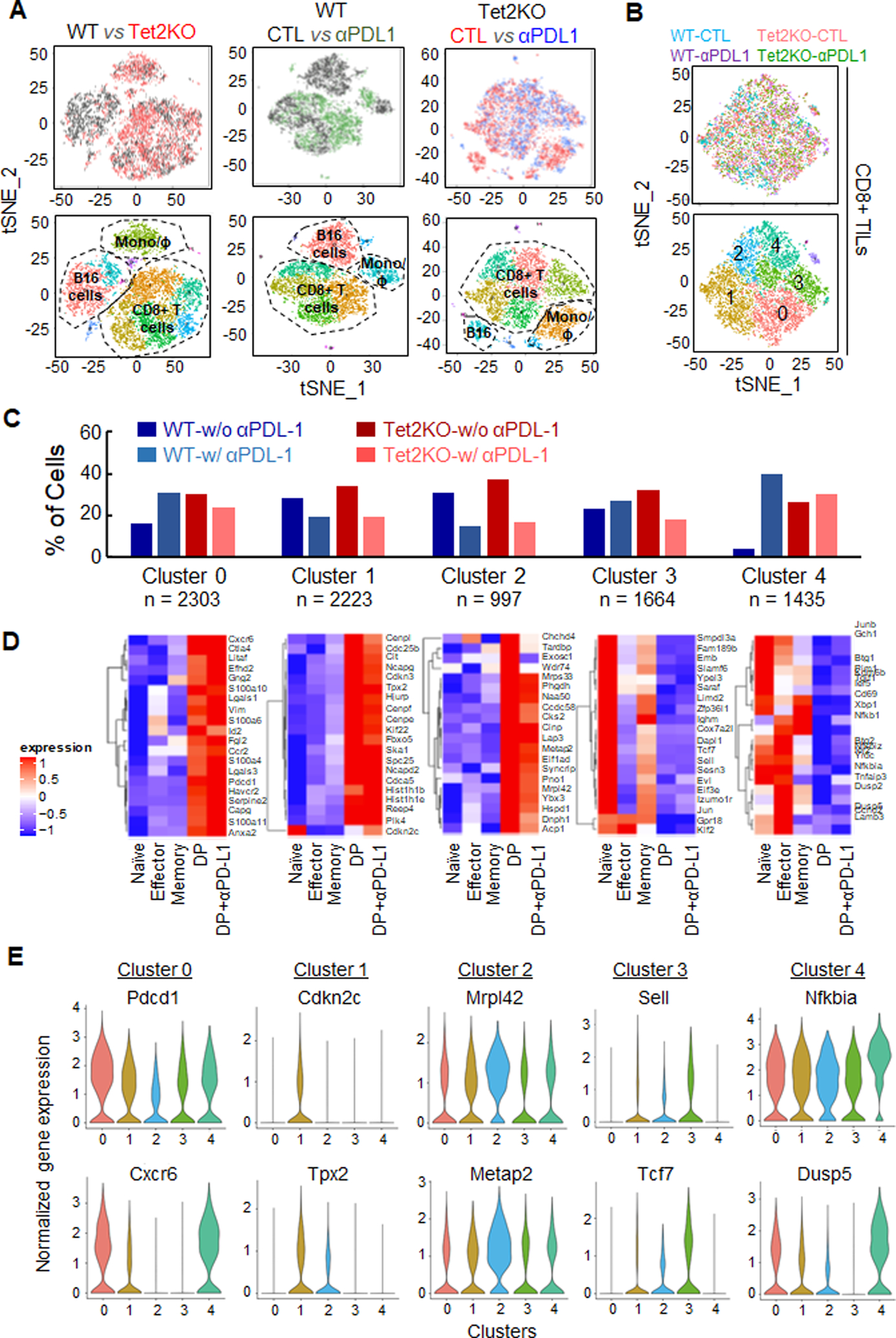
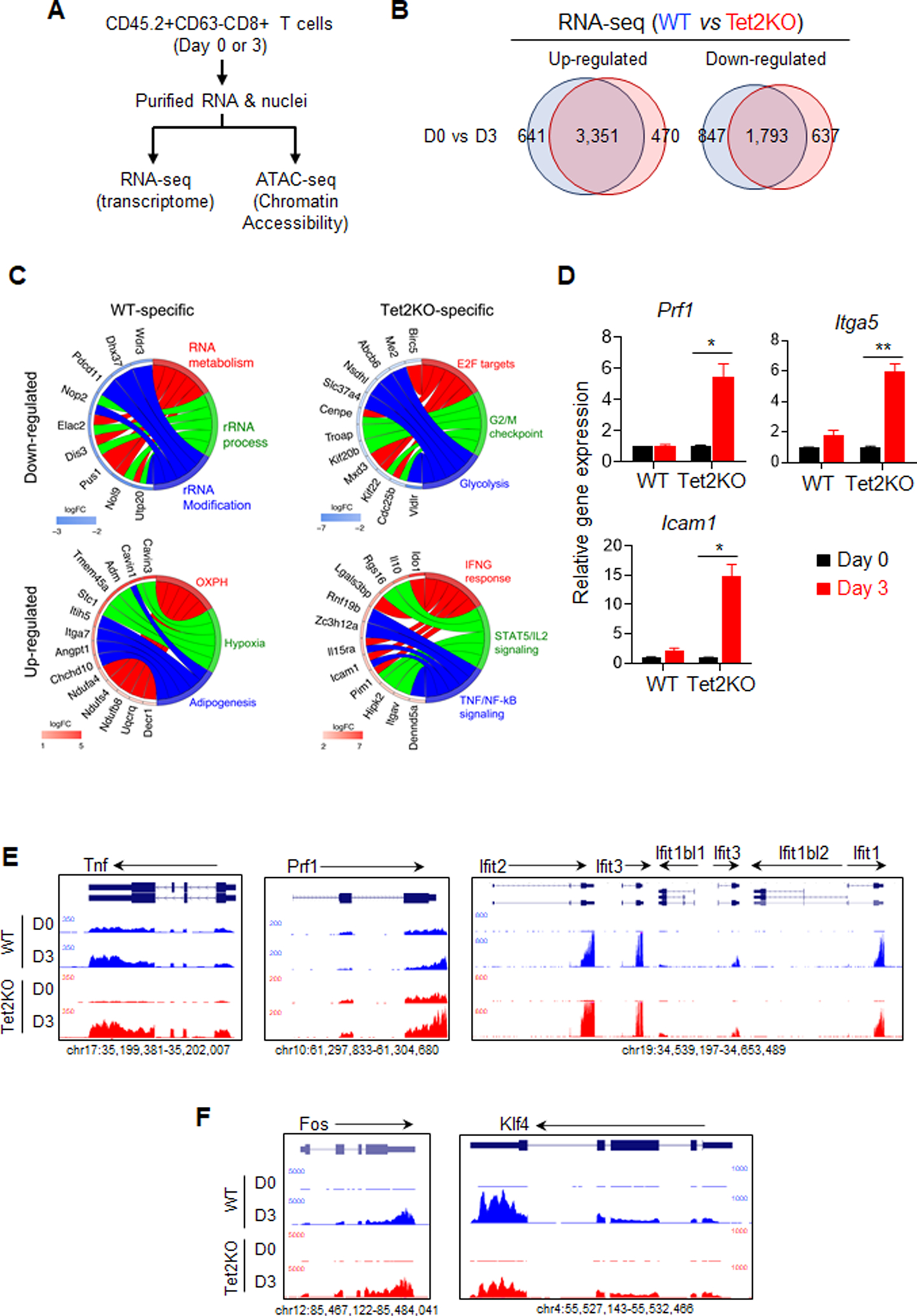
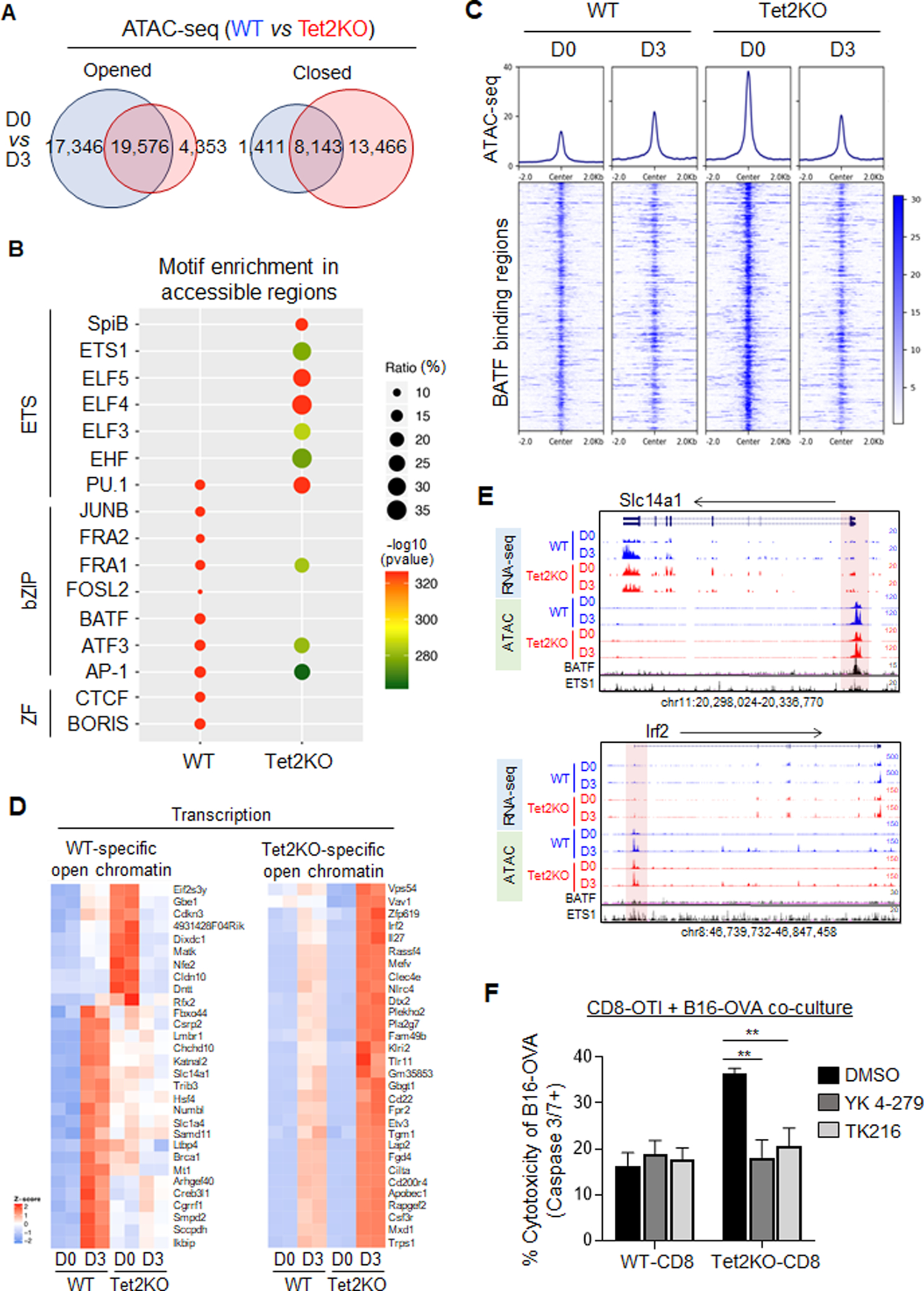
Inactivation of tumor-infiltrating lymphocytes (TIL) is one of the mechanisms mitigating antitumor immunity during tumor onset and progression. Epigenetic abnormalities are regarded as a major culprit contributing to the dysfunction of TILs within tumor microenvironments. In this study, we used a murine model of melanoma to discover that Tet2 inactivation significantly enhances the antitumor activity of TILs with an efficacy comparable to immune checkpoint inhibition imposed by anti-PD-L1 treatment. Single-cell RNA-sequencing analysis suggested that Tet2-deficient TILs exhibit effector-like features. Transcriptomic and ATAC-sequencing analysis showed that Tet2 ablation reshapes chromatin accessibility and favors binding of transcription factors geared toward CD8+ T-cell activation. Furthermore, the ETS family of transcription factors contributed to augmented CD8+ T-cell function following Tet2 depletion. Overall, our study establishes that Tet2 constitutes one of the epigenetic barriers that account for dysfunction of TILs and that Tet2 inactivation could promote antitumor immunity to suppress tumor growth. SIGNIFICANCE: This study suggests that ablation of TET2+ from TILs could promote their antitumor function by reshaping chromatin accessibility for key transcription factors and enhancing the transcription of genes essential for antitumor activity.
©2021 American Association for Cancer Research.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
