

Interspecies bacterial competition regulates community assembly in the C. elegans intestine
- PMID: 33589765
- PMCID: PMC8245486
- DOI: 10.1038/s41396-021-00910-4
Interspecies bacterial competition regulates community assembly in the C. elegans intestine
Abstract
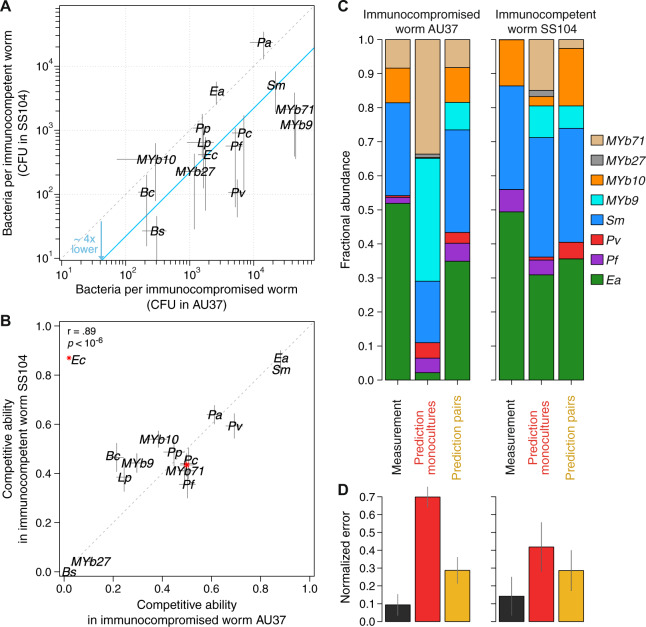
From insects to mammals, a large variety of animals hold in their intestines complex bacterial communities that play an important role in health and disease. To further our understanding of how intestinal bacterial communities assemble and function, we study the C. elegans microbiota with a bottom-up approach by feeding this nematode with bacterial monocultures as well as mixtures of two to eight bacterial species. We find that bacteria colonizing well in monoculture do not always do well in co-cultures due to interspecies bacterial interactions. Moreover, as community diversity increases, the ability to colonize the worm gut in monoculture becomes less important than interspecies interactions for determining community assembly. To explore the role of host-microbe adaptation, we compare bacteria isolated from C. elegans intestines and non-native isolates, and we find that the success of colonization is determined more by a species' taxonomy than by the isolation source. Lastly, by comparing the assembled microbiotas in two C. elegans mutants, we find that innate immunity via the p38 MAPK pathway decreases bacterial abundances yet has little influence on microbiota composition. These results highlight that bacterial interspecies interactions, more so than host-microbe adaptation or gut environmental filtering, play a dominant role in the assembly of the C. elegans microbiota.
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Nicholson JK, Holmes E, Kinross J, Burcelin R, Gibson G, Jia W, et al. Host-gut microbiota metabolic interactions. Science. 2012;336:1262–7. - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
