

Dynamics of Ku and bacterial non-homologous end-joining characterized using single DNA molecule analysis
- PMID: 33590005
- PMCID: PMC7969030
- DOI: 10.1093/nar/gkab083
Dynamics of Ku and bacterial non-homologous end-joining characterized using single DNA molecule analysis
Abstract
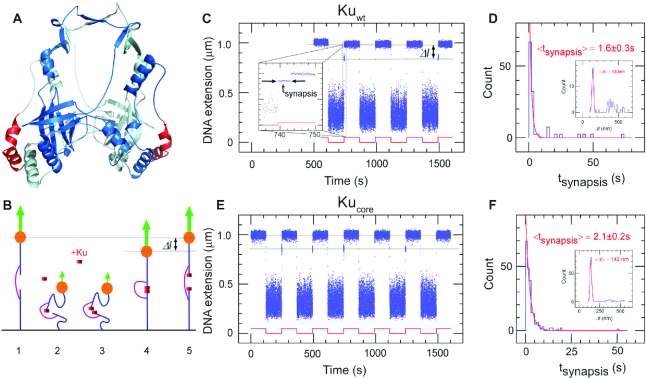
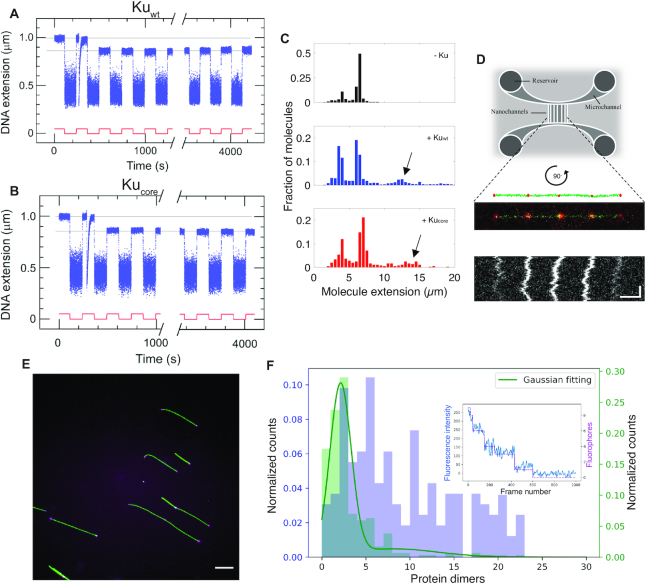
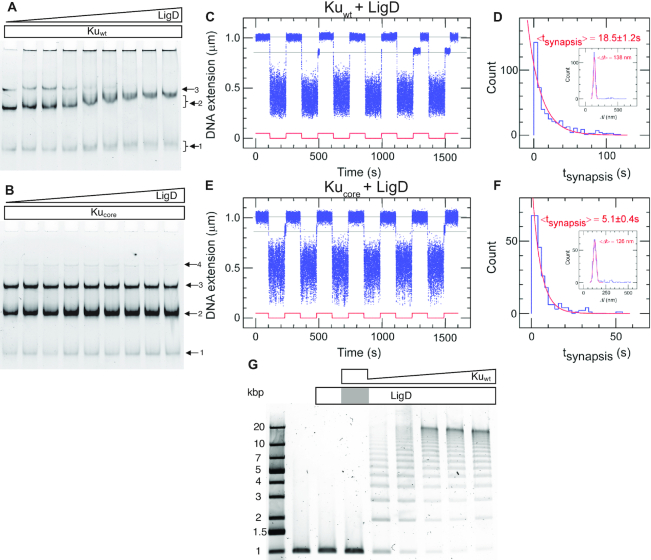
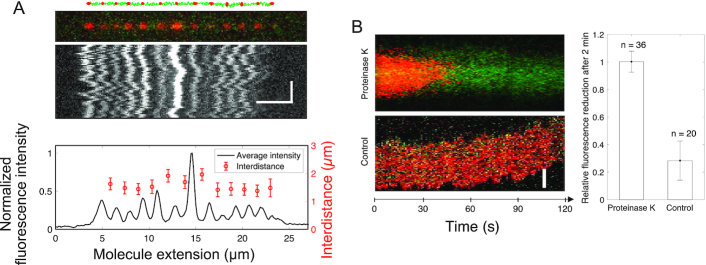
We use single-molecule techniques to characterize the dynamics of prokaryotic DNA repair by non-homologous end-joining (NHEJ), a system comprised only of the dimeric Ku and Ligase D (LigD). The Ku homodimer alone forms a ∼2 s synapsis between blunt DNA ends that is increased to ∼18 s upon addition of LigD, in a manner dependent on the C-terminal arms of Ku. The synapsis lifetime increases drastically for 4 nt complementary DNA overhangs, independently of the C-terminal arms of Ku. These observations are in contrast to human Ku, which is unable to bridge either of the two DNA substrates. We also demonstrate that bacterial Ku binds the DNA ends in a cooperative manner for synapsis initiation and remains stably bound at DNA junctions for several hours after ligation is completed, indicating that a system for removal of the proteins is active in vivo. Together these experiments shed light on the dynamics of bacterial NHEJ in DNA end recognition and processing. We speculate on the evolutionary similarities between bacterial and eukaryotic NHEJ and discuss how an increased understanding of bacterial NHEJ can open the door for future antibiotic therapies targeting this mechanism.
© The Author(s) 2021. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Shuman S., Glickman M.S.. Bacterial DNA repair by non-homologous end joining. Nat. Rev. Microbiol. 2007; 5:852–861. - PubMed
-
- Chaplin A.K., Blundell T.L.. Structural biology of multicomponent assemblies in DNA double-strand-break repair through non-homologous end joining. Curr. Opin. Struct. Biol. 2020; 61:9–16. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
