

Adaptive physiological water conservation explains hypertension and muscle catabolism in experimental chronic renal failure
- PMID: 33590667
- PMCID: PMC8244025
- DOI: 10.1111/apha.13629
Adaptive physiological water conservation explains hypertension and muscle catabolism in experimental chronic renal failure
Abstract
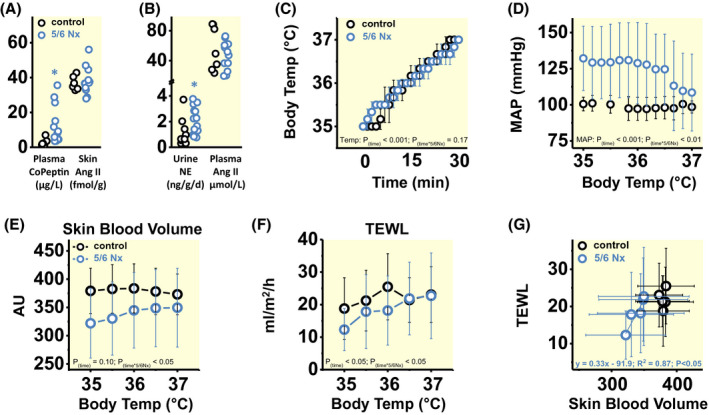
Aim: We have reported earlier that a high salt intake triggered an aestivation-like natriuretic-ureotelic body water conservation response that lowered muscle mass and increased blood pressure. Here, we tested the hypothesis that a similar adaptive water conservation response occurs in experimental chronic renal failure.
Methods: In four subsequent experiments in Sprague Dawley rats, we used surgical 5/6 renal mass reduction (5/6 Nx) to induce chronic renal failure. We studied solute and water excretion in 24-hour metabolic cage experiments, chronic blood pressure by radiotelemetry, chronic metabolic adjustment in liver and skeletal muscle by metabolomics and selected enzyme activity measurements, body Na+ , K+ and water by dry ashing, and acute transepidermal water loss in conjunction with skin blood flow and intra-arterial blood pressure.
Results: 5/6 Nx rats were polyuric, because their kidneys could not sufficiently concentrate the urine. Physiological adaptation to this renal water loss included mobilization of nitrogen and energy from muscle for organic osmolyte production, elevated norepinephrine and copeptin levels with reduced skin blood flow, which by means of compensation reduced their transepidermal water loss. This complex physiologic-metabolic adjustment across multiple organs allowed the rats to stabilize their body water content despite persisting renal water loss, albeit at the expense of hypertension and catabolic mobilization of muscle protein.
Conclusion: Physiological adaptation to body water loss, termed aestivation, is an evolutionary conserved survival strategy and an under-studied research area in medical physiology, which besides hypertension and muscle mass loss in chronic renal failure may explain many otherwise unexplainable phenomena in medicine.
Keywords: aestivation; body sodium; body water; dehydration; double-barrier concept; glucose-alanine-shuttle; glycine methylation; hepato-renal; hypertension; kidney; liver; muscle mass loss; organic osmolytes; purine metabolism; skin; transamination; transepidermal water loss; urea cycle; urine concentration.
© 2021 The Authors. Acta Physiologica published by John Wiley & Sons Ltd on behalf of Scandinavian Physiological Society.
Conflict of interest statement
The authors declare no competing interests.
Figures




Comment in
-
Hypertension due to loss of water.Acta Physiol (Oxf). 2021 Jun;232(2):e13658. doi: 10.1111/apha.13658. Epub 2021 Apr 18. Acta Physiol (Oxf). 2021. PMID: 33819385 No abstract available.
References
-
- Borst JG, Borst‐De GA. Hypertension explained by Starling's theory of circulatory homoeostasis. Lancet. 1963;1(7283):677‐682. - PubMed
-
- Coleman TG, Guyton AC. Hypertension caused by salt loading in the dog. 3. Onset transients of cardiac output and other circulatory variables. Circ Res. 1969;25(2):153‐160. - PubMed
-
- Guyton AC, Coleman TG, Cowley AWJ, Manning RDJ, Norman RA, Ferguson JD. Brief reviews: a systems analysis approach to understanding long‐range arterial blood pressure control and hypertension. Circ Res. 1974;35:159‐176.
-
- Guyton AC. Blood pressure control–special role of the kidneys and body fluids. Science. 1991;252(5014):1813‐1816. - PubMed
-
- Montani JP, Van Vliet BN. Understanding the contribution of Guyton's large circulatory model to long‐term control of arterial pressure. Exp Physiol. 2009;94(4):382‐388. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
