

Non-permissive human conventional CD1c+ dendritic cells enable trans-infection of human primary renal tubular epithelial cells and protect BK polyomavirus from neutralization
- PMID: 33592065
- PMCID: PMC7886149
- DOI: 10.1371/journal.ppat.1009042
Non-permissive human conventional CD1c+ dendritic cells enable trans-infection of human primary renal tubular epithelial cells and protect BK polyomavirus from neutralization
Abstract
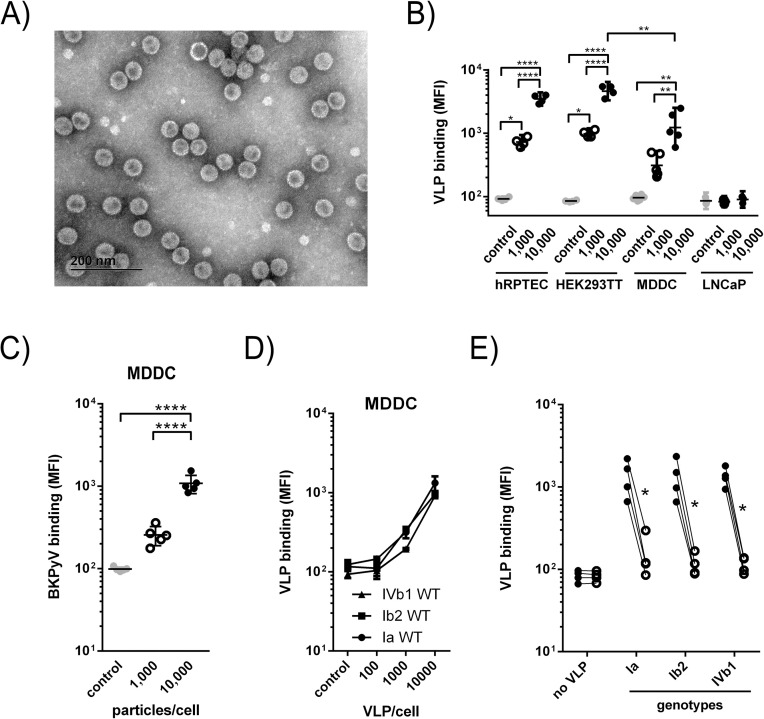
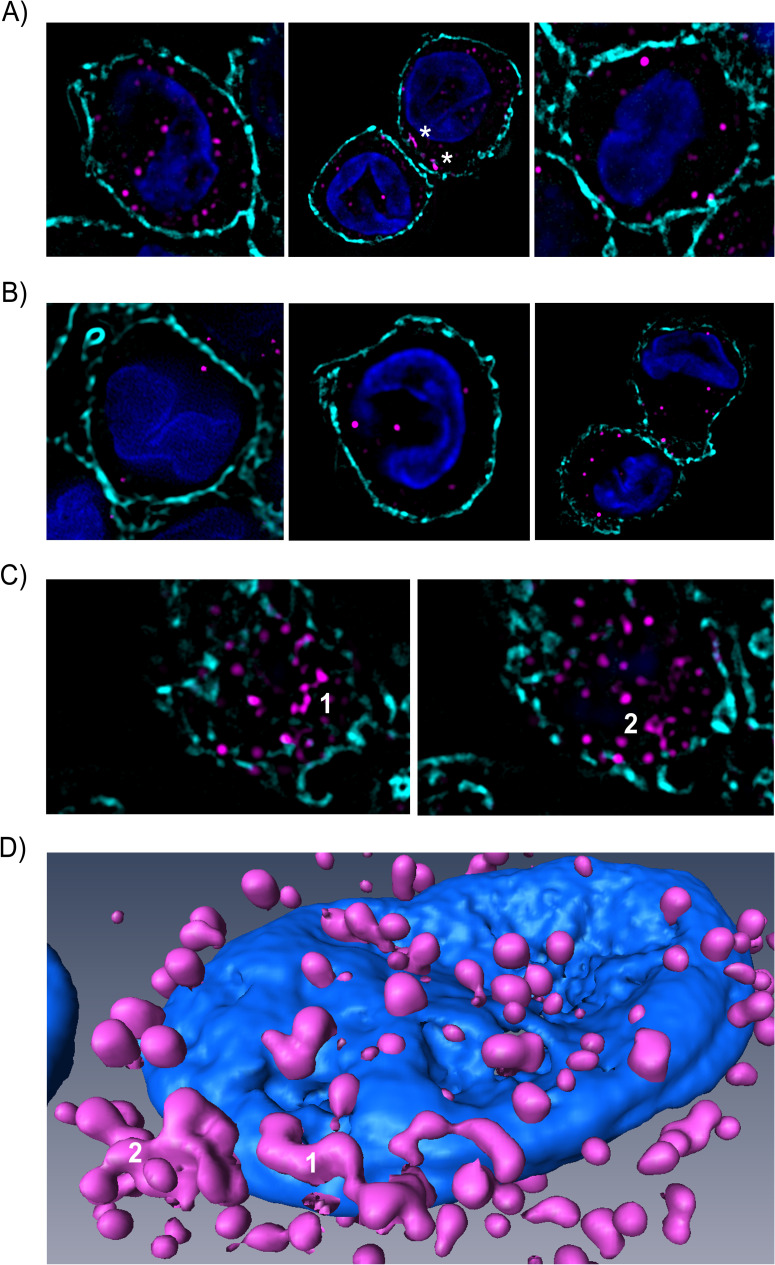
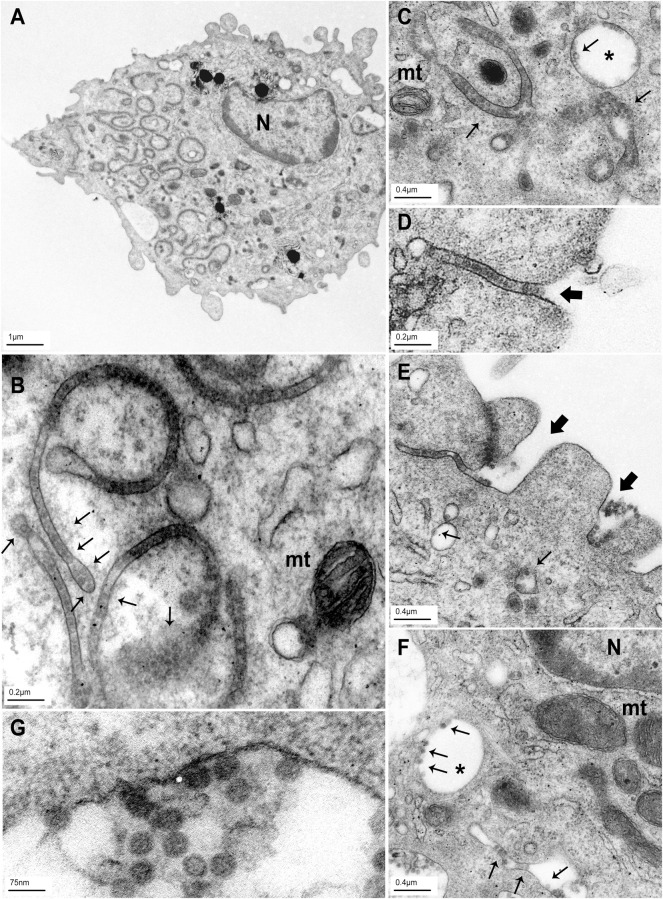
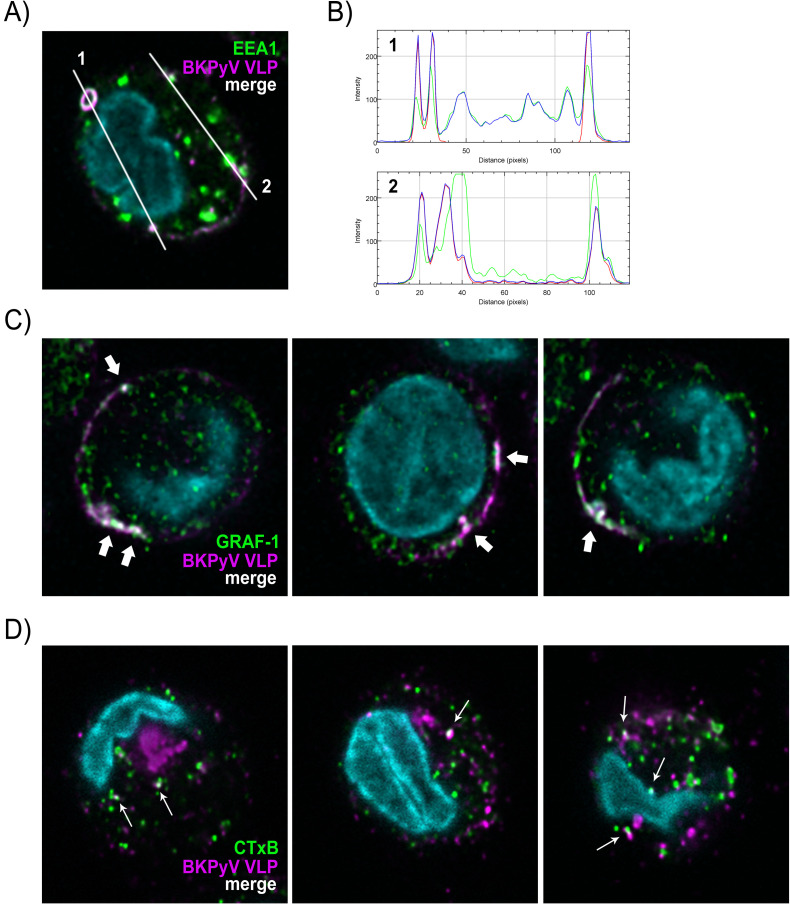
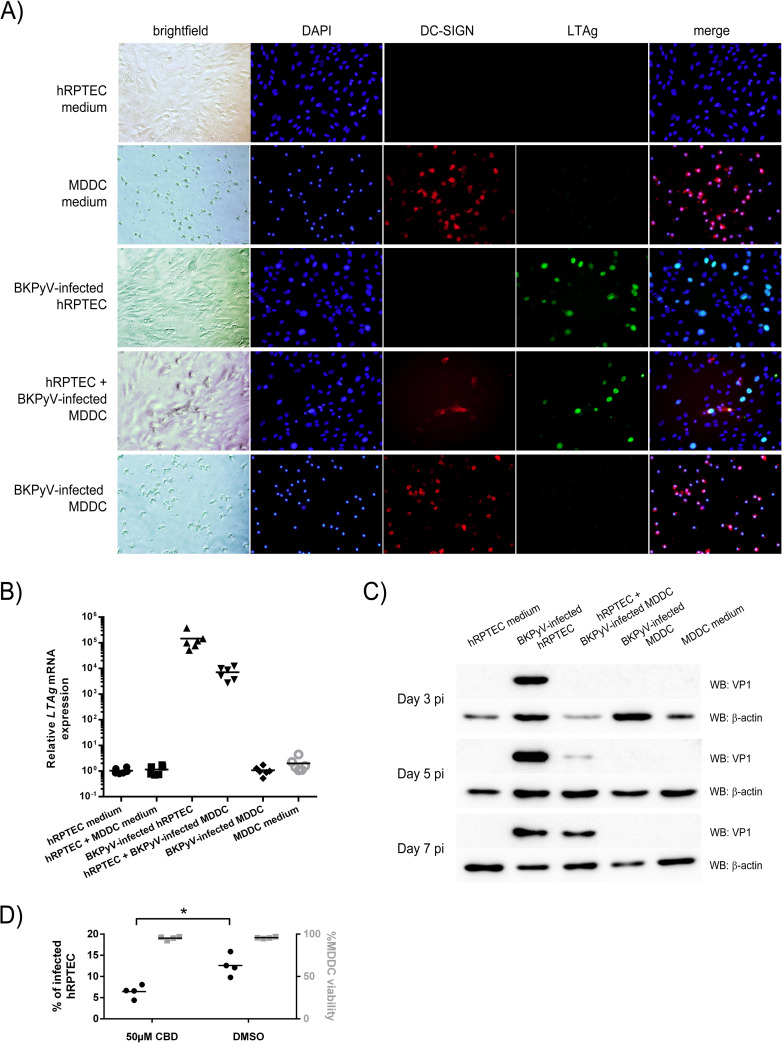
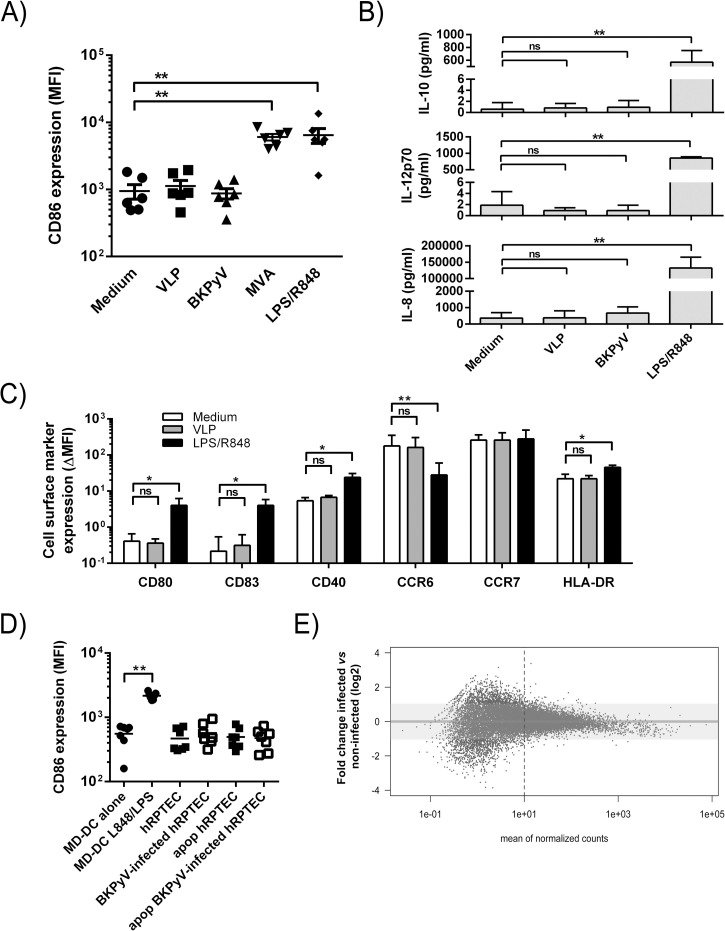
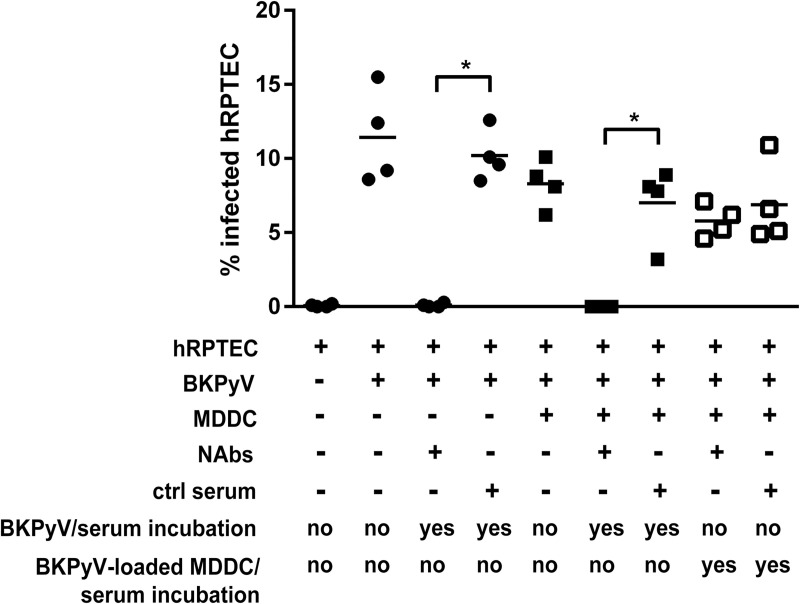
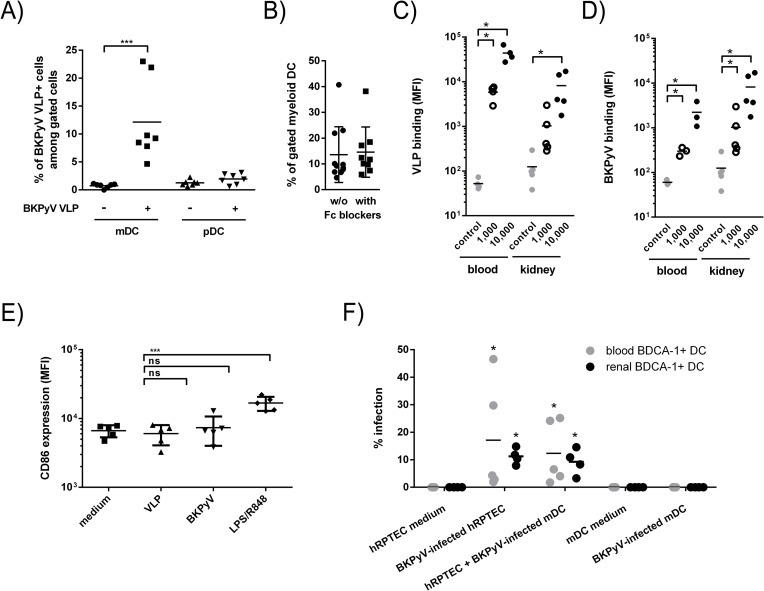
The BK polyomavirus (BKPyV) is a ubiquitous human virus that persists in the renourinary epithelium. Immunosuppression can lead to BKPyV reactivation in the first year post-transplantation in kidney transplant recipients (KTRs) and hematopoietic stem cell transplant recipients. In KTRs, persistent DNAemia has been correlated to the occurrence of polyomavirus-associated nephropathy (PVAN) that can lead to graft loss if not properly controlled. Based on recent observations that conventional dendritic cells (cDCs) specifically infiltrate PVAN lesions, we hypothesized that those cells could play a role in BKPyV infection. We first demonstrated that monocyte-derived dendritic cells (MDDCs), an in vitro model for mDCs, captured BKPyV particles through an unconventional GRAF-1 endocytic pathway. Neither BKPyV particles nor BKPyV-infected cells were shown to activate MDDCs. Endocytosed virions were efficiently transmitted to permissive cells and protected from the antibody-mediated neutralization. Finally, we demonstrated that freshly isolated CD1c+ mDCs from the blood and kidney parenchyma behaved similarly to MDDCs thus extending our results to cells of clinical relevance. This study sheds light on a potential unprecedented CD1c+ mDC involvement in the BKPyV infection as a promoter of viral spreading.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
BK polyomavirus serotype-specific antibody responses in blood donors and kidney transplant recipients with and without new-onset BK polyomavirus-DNAemia: A Swiss Transplant Cohort Study.Am J Transplant. 2025 May;25(5):985-1001. doi: 10.1016/j.ajt.2024.11.019. Epub 2024 Nov 22. Am J Transplant. 2025. PMID: 39580075
-
BK Polyomavirus-specific T Cells as a Diagnostic and Prognostic Marker for BK Polyomavirus Infections After Pediatric Kidney Transplantation.Transplantation. 2020 Nov;104(11):2393-2402. doi: 10.1097/TP.0000000000003133. Transplantation. 2020. PMID: 31985731
-
Stability of BK polyomavirus IgG seroreactivity and its correlation with preceding viremia.J Clin Virol. 2017 May;90:46-51. doi: 10.1016/j.jcv.2017.03.015. Epub 2017 Mar 19. J Clin Virol. 2017. PMID: 28355579
-
Diagnostics, treatment, and immune response in BK polyomavirus infection after pediatric kidney transplantation.Pediatr Nephrol. 2020 Mar;35(3):375-382. doi: 10.1007/s00467-018-4164-3. Epub 2018 Dec 11. Pediatr Nephrol. 2020. PMID: 30539254 Review.
-
The Second International Consensus Guidelines on the Management of BK Polyomavirus in Kidney Transplantation.Transplantation. 2024 Sep 1;108(9):1834-1866. doi: 10.1097/TP.0000000000004976. Epub 2024 Apr 12. Transplantation. 2024. PMID: 38605438 Free PMC article. Review.
Cited by
-
Neutralizing Antibodies Targeting BK Polyomavirus: Clinical Importance and Therapeutic Potential for Kidney Transplant Recipients.J Am Soc Nephrol. 2024 Oct 1;35(10):1425-1433. doi: 10.1681/ASN.0000000000000457. Epub 2024 Jul 9. J Am Soc Nephrol. 2024. PMID: 39352862 Review.
-
Digital spatial profiling reveals the molecular signatures of BK virus infection in renal transplant recipients.Transl Androl Urol. 2025 Jul 30;14(7):2089-2105. doi: 10.21037/tau-2025-451. Epub 2025 Jul 28. Transl Androl Urol. 2025. PMID: 40800081 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
