

A cold-stress-inducible PERK/OGT axis controls TOM70-assisted mitochondrial protein import and cristae formation
- PMID: 33592173
- PMCID: PMC7962155
- DOI: 10.1016/j.cmet.2021.01.013
A cold-stress-inducible PERK/OGT axis controls TOM70-assisted mitochondrial protein import and cristae formation
Abstract
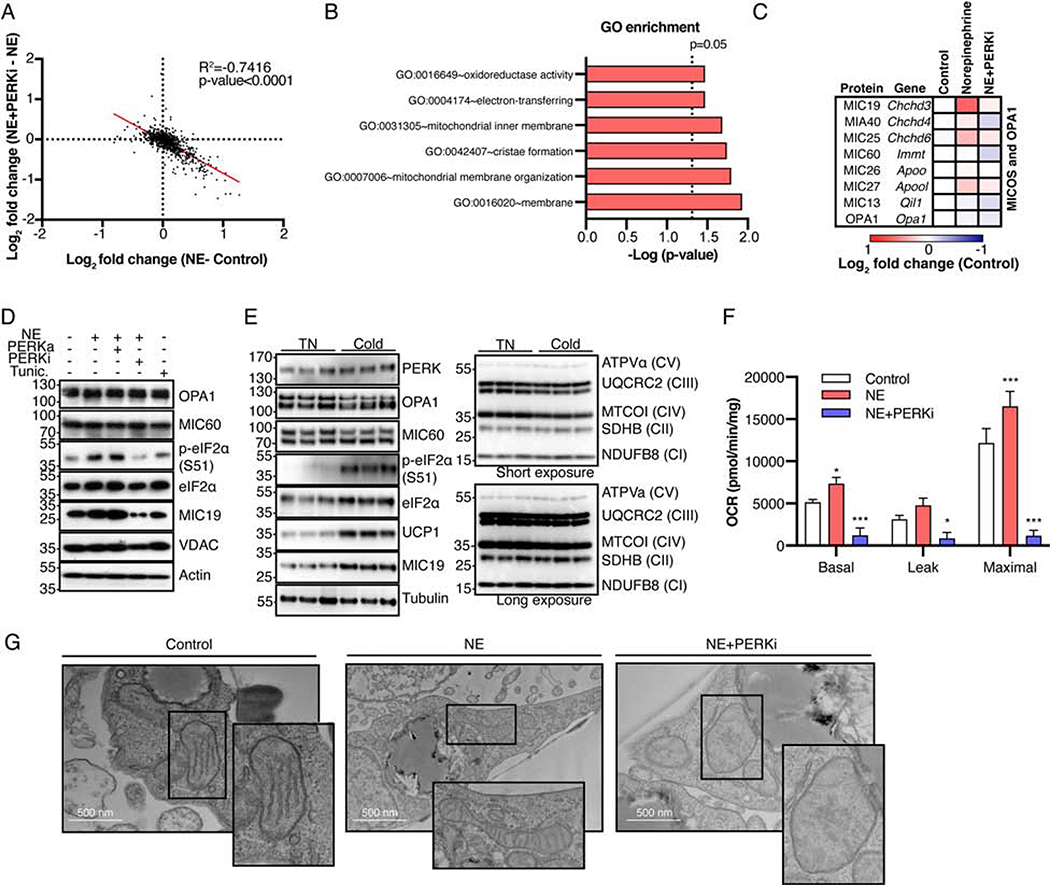
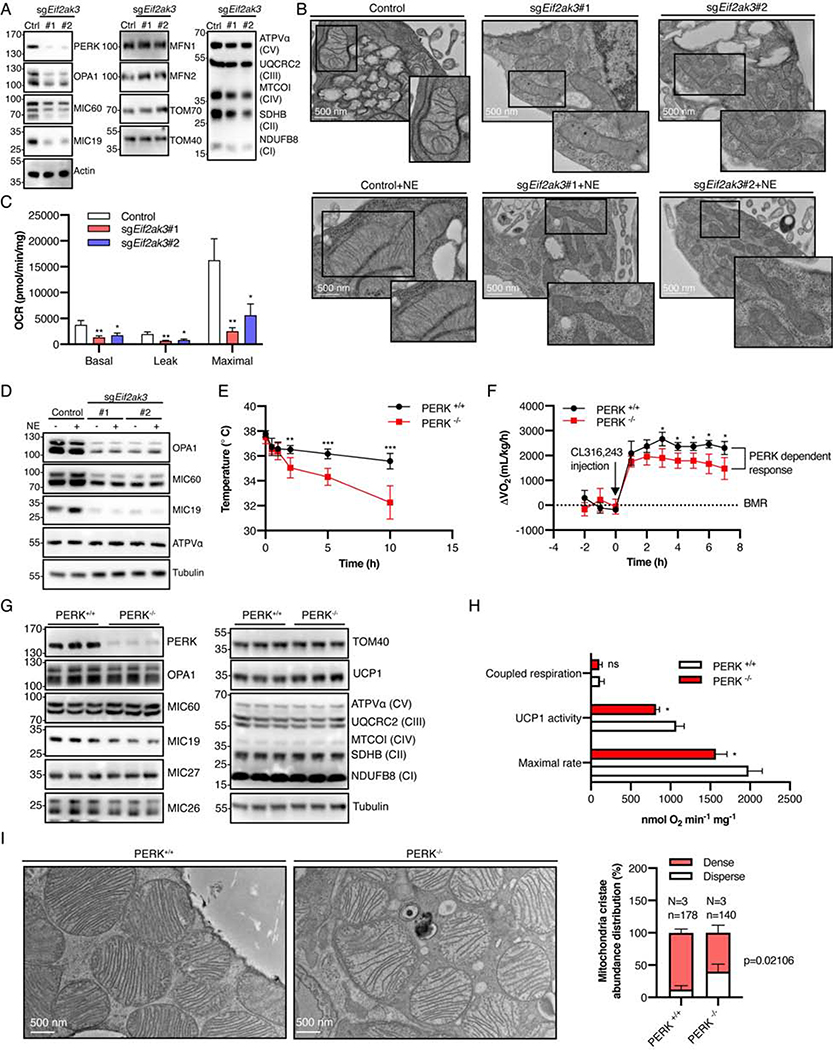
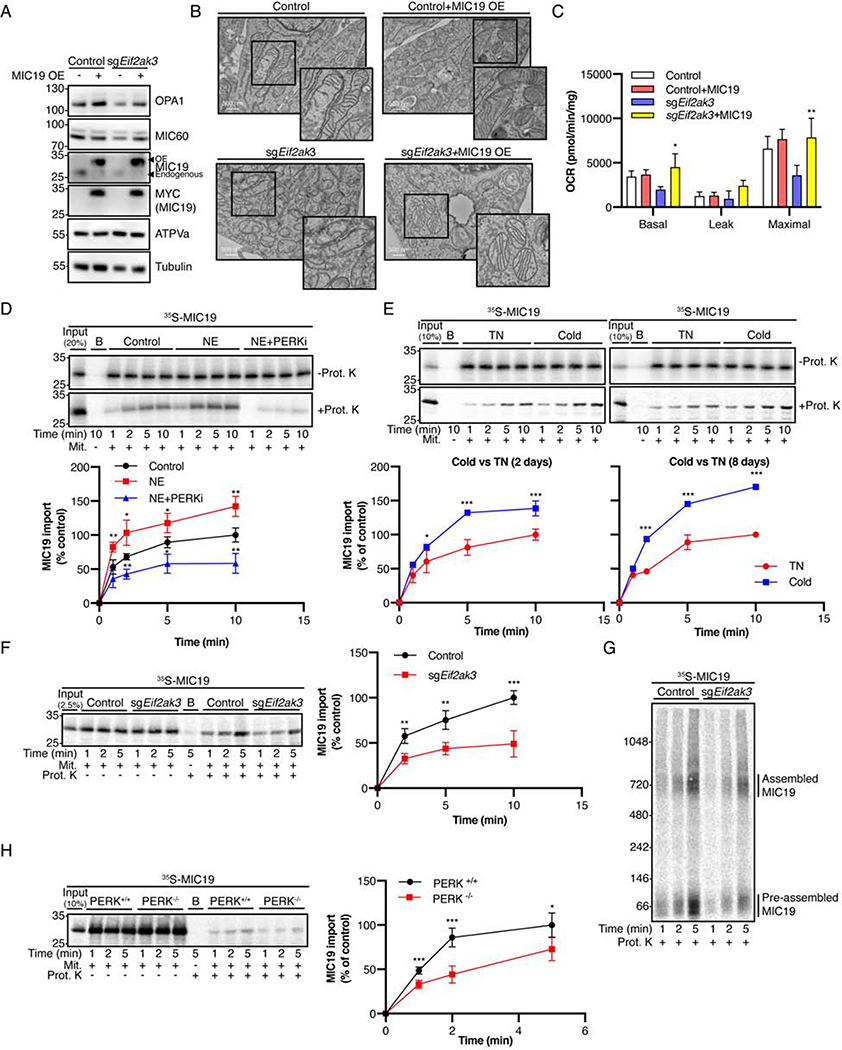
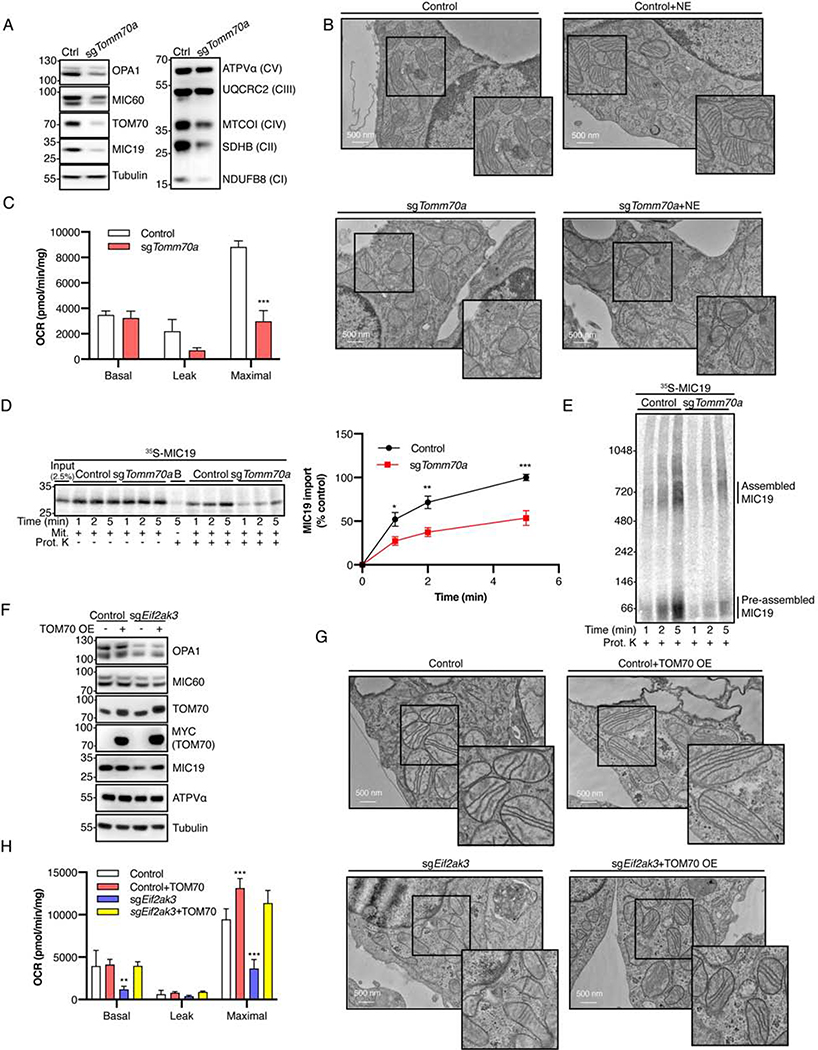
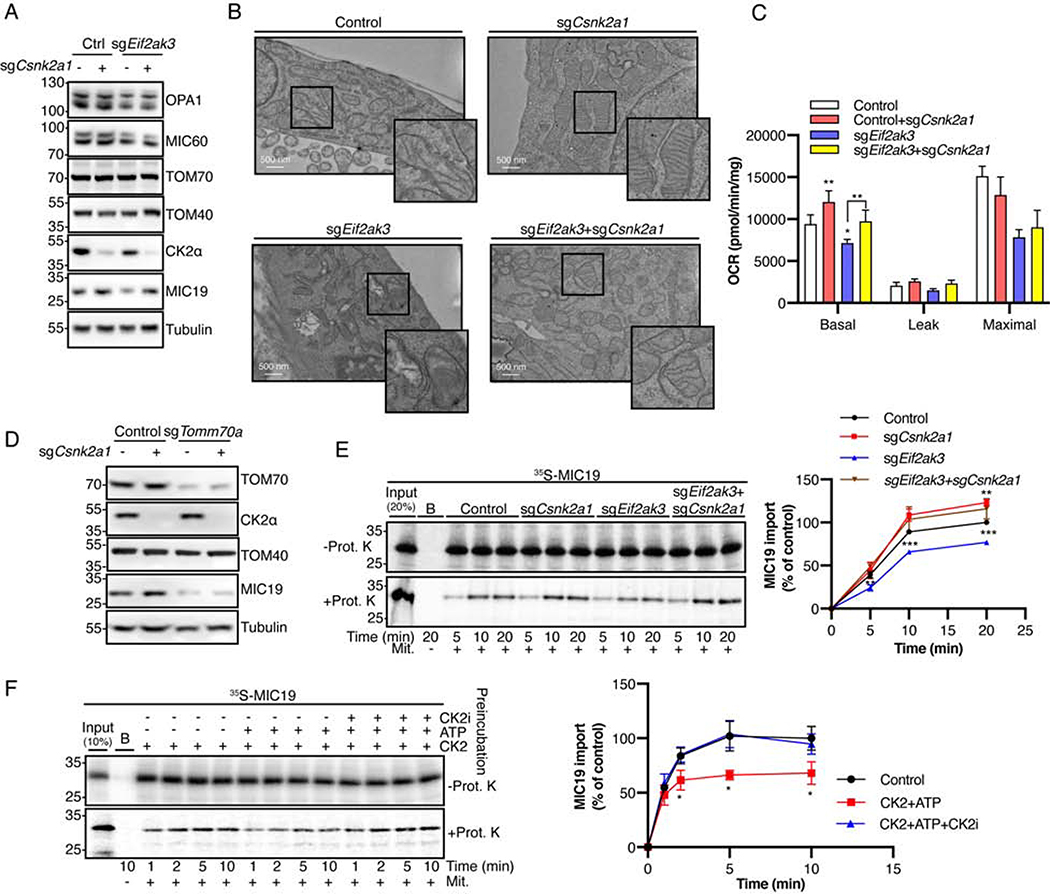
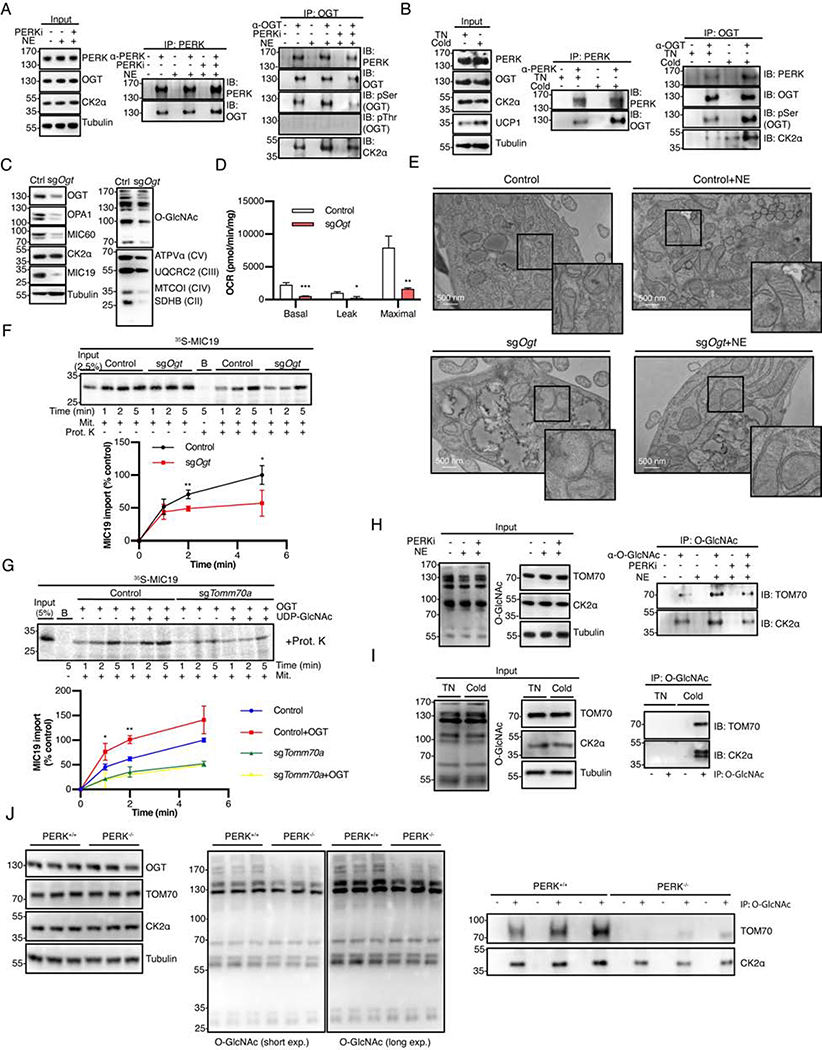
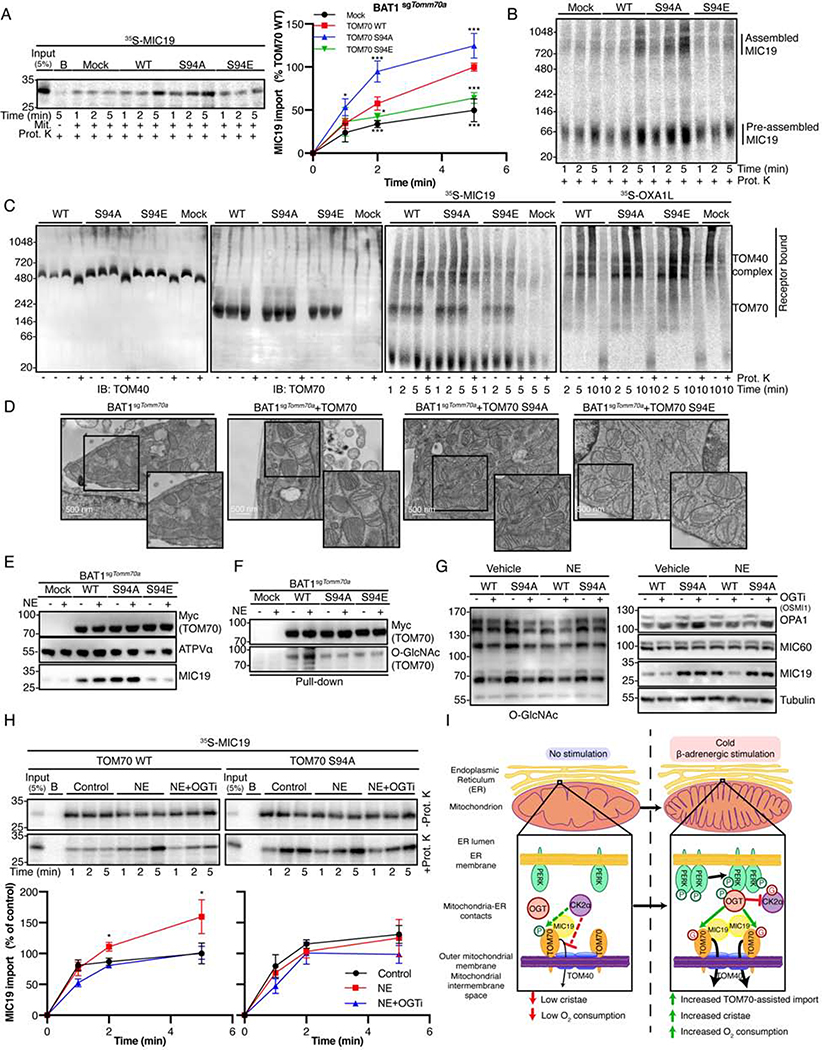
The architecture of cristae provides a spatial mitochondrial organization that contains functional respiratory complexes. Several protein components including OPA1 and MICOS complex subunits organize cristae structure, but upstream regulatory mechanisms are largely unknown. Here, in vivo and in vitro reconstitution experiments show that the endoplasmic reticulum (ER) kinase PERK promotes cristae formation by increasing TOM70-assisted mitochondrial import of MIC19, a critical subunit of the MICOS complex. Cold stress or β-adrenergic stimulation activates PERK that phosphorylates O-linked N-acetylglucosamine transferase (OGT). Phosphorylated OGT glycosylates TOM70 on Ser94, enhancing MIC19 protein import into mitochondria and promoting cristae formation and respiration. In addition, PERK-activated OGT O-GlcNAcylates and attenuates CK2α activity, which mediates TOM70 Ser94 phosphorylation and decreases MIC19 mitochondrial protein import. We have identified a cold-stress inter-organelle PERK-OGT-TOM70 axis that increases cell respiration through mitochondrial protein import and subsequent cristae formation. These studies have significant implications in cellular bioenergetics and adaptations to stress conditions.
Keywords: MIC19; PERK-OGT axis; TOM70; brown adipocytes; cold stress; cristae biogenesis; mitochondrial protein import; respiration.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Abe Y, Shodai T, Muto T, Mihara K, Torii H, Nishikawa S. ichi, Endo T, and Kohda D (2000). Structural basis of presequence recognition by the mitochondrial protein import receptor Tom20. Cell 100, 551–560. - PubMed
-
- Akabane S, Uno M, Tani N, Shimazaki S, Ebara N, Kato H, Kosako H, and Oka T (2016). PKA Regulates PINK1 Stability and Parkin Recruitment to Damaged Mitochondria through Phosphorylation of MIC60. Mol. Cell 62, 371–384. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
