

ERβ regulated ovarian kisspeptin plays an important role in oocyte maturation
- PMID: 33592287
- PMCID: PMC8906370
- DOI: 10.1016/j.mce.2021.111208
ERβ regulated ovarian kisspeptin plays an important role in oocyte maturation
Abstract
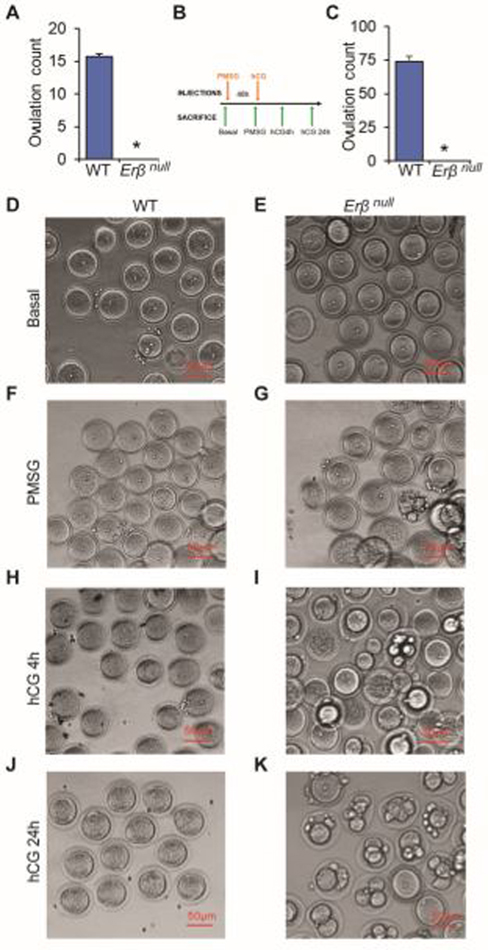
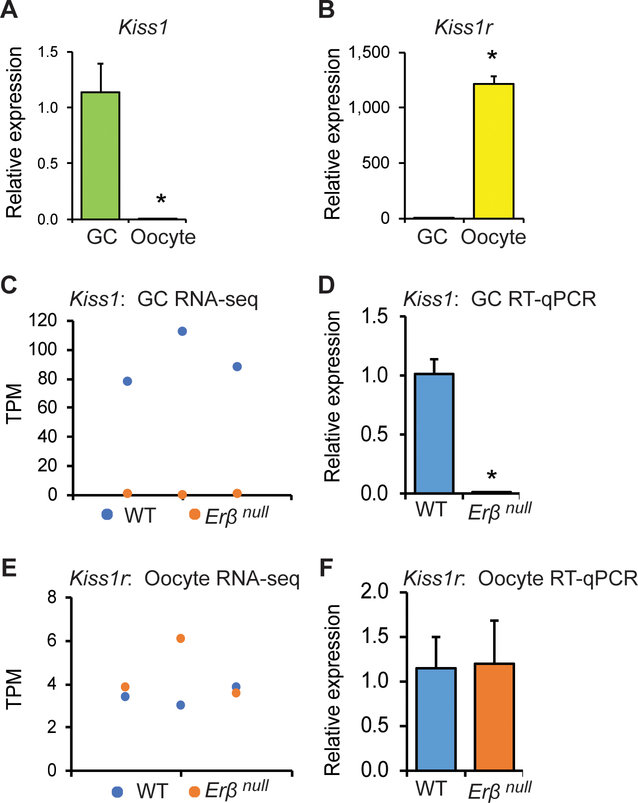
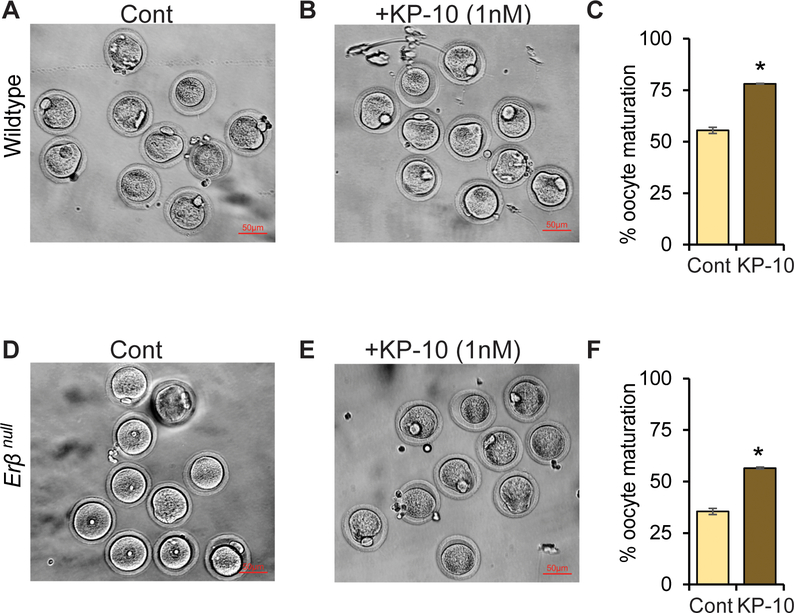
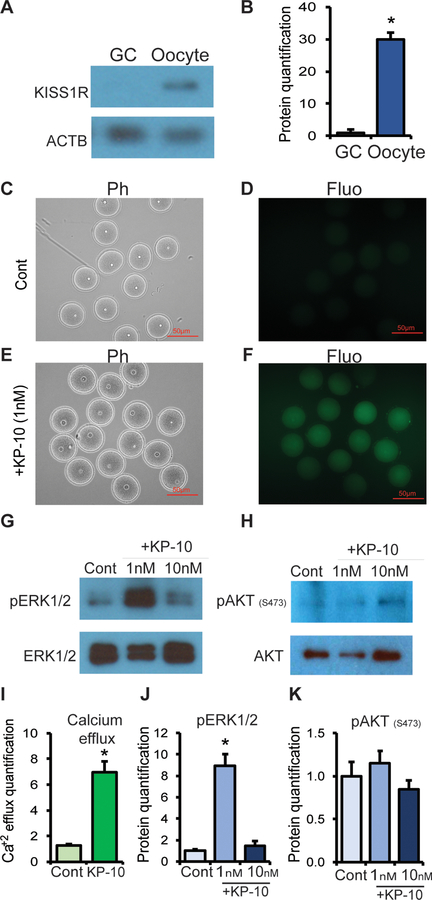
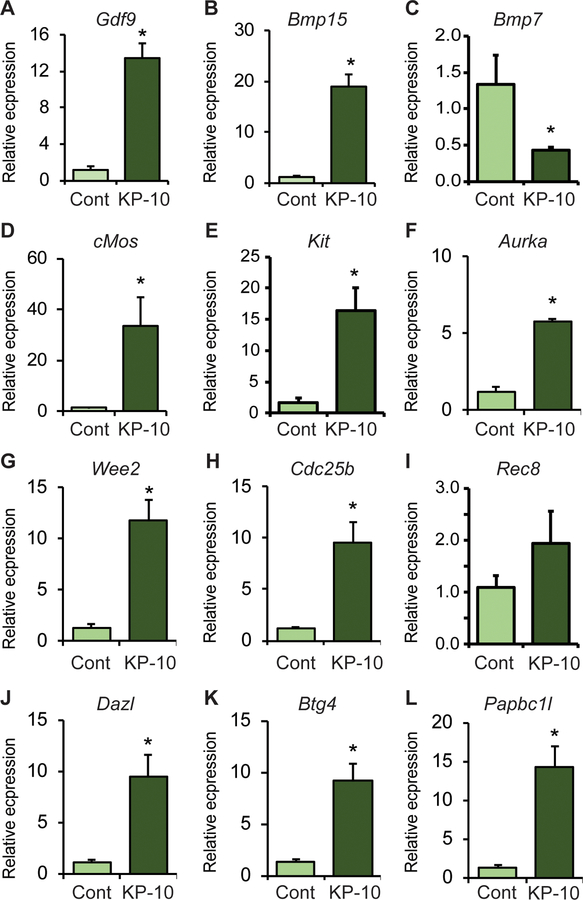
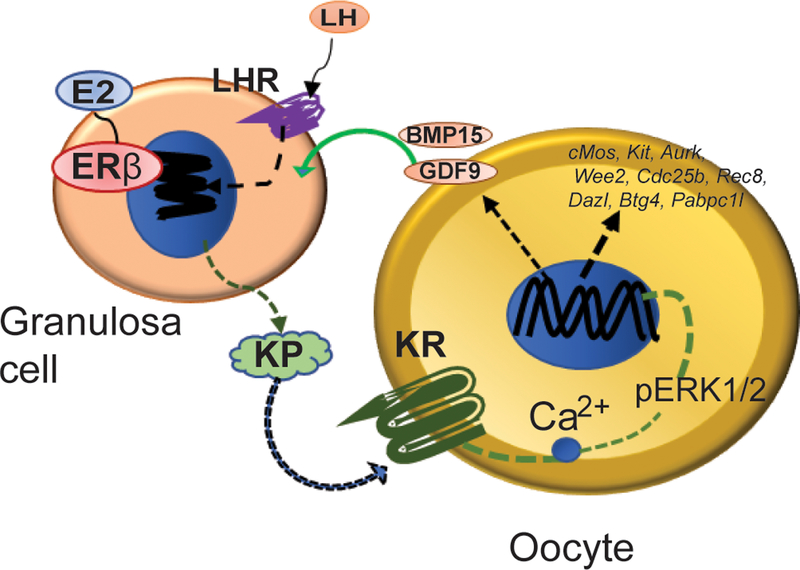
Kisspeptin (KISS1) signaling in the hypothalamic-pituitary (H-P) axis plays an essential role in regulating gonadotropin secretion. KISS1 and KISS1 receptor (KISS1R) are also expressed in the ovary; however, the role of intraovarian KISS1 signaling remains unclear. Granulosa cell (GC)-specific expression of KISS1, and oocyte-specific expression of KISS1R indicate that GC-derived KISS1 may act on oocytes. Expression of KISS1 in GCs is induced by gonadotropins but it is absent in estrogen receptor β knockout (Erβnull) rat ovaries. We also observed that gonadotropin stimulation failed to induce maturation of Erβnull oocytes. Interestingly, KISS1 treatment of cumulus oocyte complexes (COCs) isolated from antral follicles promotes in vitro maturation of oocytes. Treatment of oocytes with KISS1 induced intracellular Ca2+ release, and increased activation of MAP kinase ERK1/2. KISS1 treatment also induced the expression of oocyte genes that are crucial for differentiation of GCs, and maturation of oocytes. Our findings suggest that ovarian KISS1-signaling plays an important role in gonadotropin induced follicle development and oocyte maturation.
Keywords: And oocyte maturation; Estrogen receptor β; Gonadotropin; Granulosa cells; Kisspeptin; Kisspeptin receptor.
Copyright © 2021. Published by Elsevier B.V.
Conflict of interest statement
Figures
Similar articles
-
ERβ Regulation of Gonadotropin Responses during Folliculogenesis.Int J Mol Sci. 2021 Sep 26;22(19):10348. doi: 10.3390/ijms221910348. Int J Mol Sci. 2021. PMID: 34638689 Free PMC article. Review.
-
ESR2 Is Essential for Gonadotropin-Induced Kiss1 Expression in Granulosa Cells.Endocrinology. 2018 Nov 1;159(11):3860-3873. doi: 10.1210/en.2018-00608. Endocrinology. 2018. PMID: 30277501 Free PMC article.
-
The Kisspeptin and Kisspeptin receptor in follicular microenvironment: is that really necessary for oocyte maturation and fertilisation?J Obstet Gynaecol. 2022 Oct;42(7):3241-3247. doi: 10.1080/01443615.2022.2112018. Epub 2022 Aug 22. J Obstet Gynaecol. 2022. PMID: 35993609
-
The role of Kisspeptin signaling in Oocyte maturation.Front Endocrinol (Lausanne). 2022 Aug 22;13:917464. doi: 10.3389/fendo.2022.917464. eCollection 2022. Front Endocrinol (Lausanne). 2022. PMID: 36072937 Free PMC article. Review.
-
Expression of neurokinin B/NK3 receptor and kisspeptin/KISS1 receptor in human granulosa cells.Hum Reprod. 2014 Dec;29(12):2736-46. doi: 10.1093/humrep/deu247. Epub 2014 Oct 14. Hum Reprod. 2014. PMID: 25316443
Cited by
-
IGF-1 Acts through Kiss1-expressing Cells to Influence Metabolism and Reproduction.bioRxiv [Preprint]. 2024 Jul 4:2024.07.02.601722. doi: 10.1101/2024.07.02.601722. bioRxiv. 2024. PMID: 39005405 Free PMC article. Preprint.
-
Bisphenol A Disrupts Ribosome Function during Ovarian Development of Mice.Toxics. 2024 Aug 26;12(9):627. doi: 10.3390/toxics12090627. Toxics. 2024. PMID: 39330555 Free PMC article.
-
Sexual Dimorphism in Kisspeptin Signaling.Cells. 2022 Mar 28;11(7):1146. doi: 10.3390/cells11071146. Cells. 2022. PMID: 35406710 Free PMC article. Review.
-
Early Development of Hypothalamic Neurons Expressing Proopiomelanocortin Peptides, Neuropeptide Y, and Kisspeptin in Fetal Rhesus Macaques.eNeuro. 2025 Jul 3;12(7):ENEURO.0087-25.2025. doi: 10.1523/ENEURO.0087-25.2025. Print 2025 Jul. eNeuro. 2025. PMID: 40389300 Free PMC article.
-
ERβ Regulation of Gonadotropin Responses during Folliculogenesis.Int J Mol Sci. 2021 Sep 26;22(19):10348. doi: 10.3390/ijms221910348. Int J Mol Sci. 2021. PMID: 34638689 Free PMC article. Review.
References
-
- Araki K, Naito K, Haraguchi S, Suzuki R, Yokoyama M, Inoue M, Aizawa S, Toyoda Y, and Sato E. 1996. Meiotic abnormalities of c-mos knockout mouse oocytes: activation after first meiosis or entrance into third meiotic metaphase. Biology of reproduction. 55:1315–1324. - PubMed
-
- Asadi M, Ghafouri-Fard S, Zare-Abdollahi D, Ebrahim-Habibi A, and Matin N. 2013. Estrogen receptor mutation in a girl with primary amenorrhea. Clinical genetics. 83:497–498. - PubMed
-
- Atherfold PA, Norris MS, Robinson PJ, Gelfand EW, and Franklin RA. 1999. Calcium-induced ERK activation in human T lymphocytes. Molecular immunology. 36:543–549. - PubMed
-
- Byers M, Kuiper GG, Gustafsson JA, and Park-Sarge OK. 1997. Estrogen receptor-beta mRNA expression in rat ovary: down-regulation by gonadotropins. Molecular endocrinology (Baltimore, Md.). 11:172–182. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
