

Diversity of Receptive Fields and Sideband Inhibition with Complex Thalamocortical and Intracortical Origin in L2/3 of Mouse Primary Auditory Cortex
- PMID: 33593857
- PMCID: PMC8026349
- DOI: 10.1523/JNEUROSCI.1732-20.2021
Diversity of Receptive Fields and Sideband Inhibition with Complex Thalamocortical and Intracortical Origin in L2/3 of Mouse Primary Auditory Cortex
Abstract
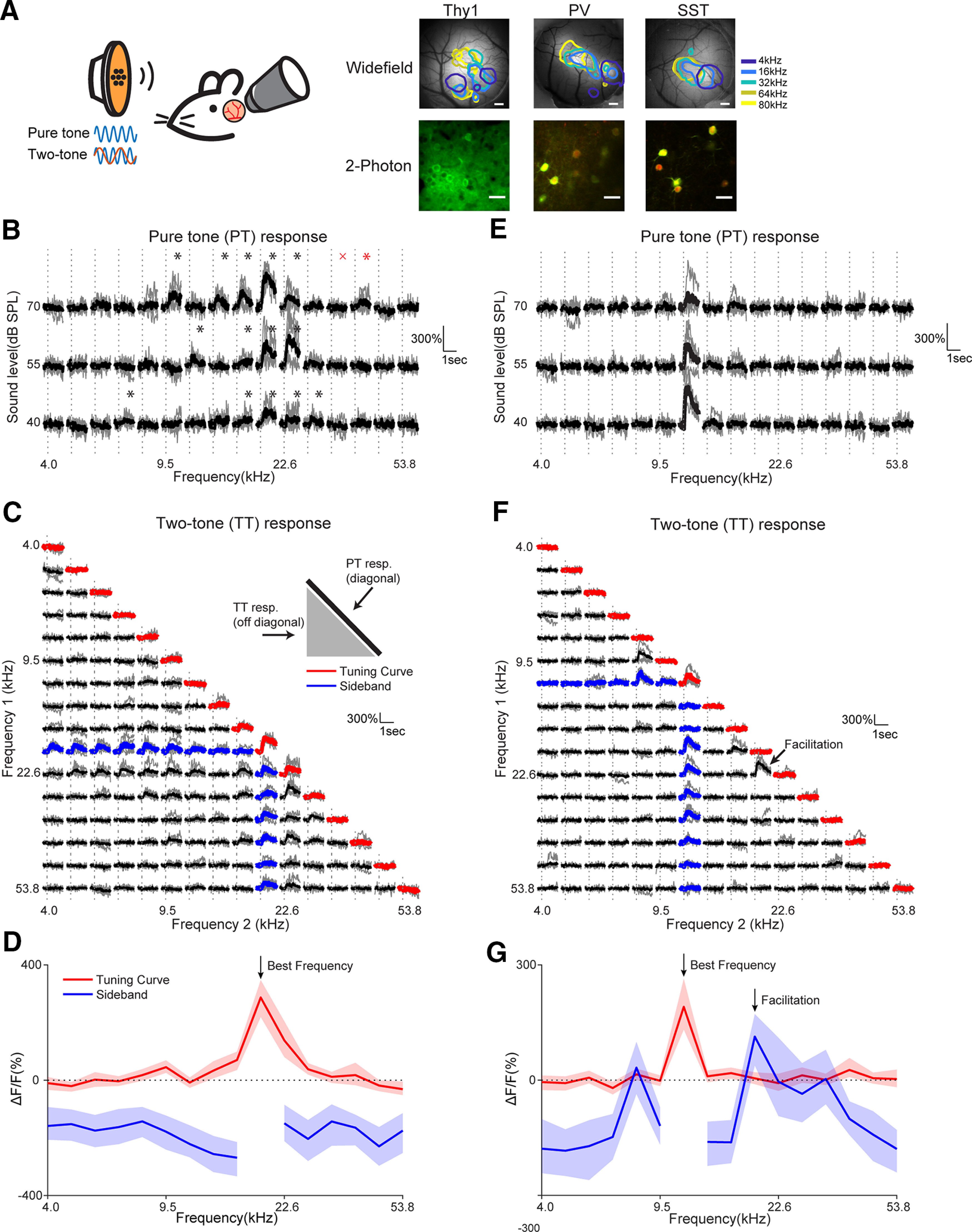
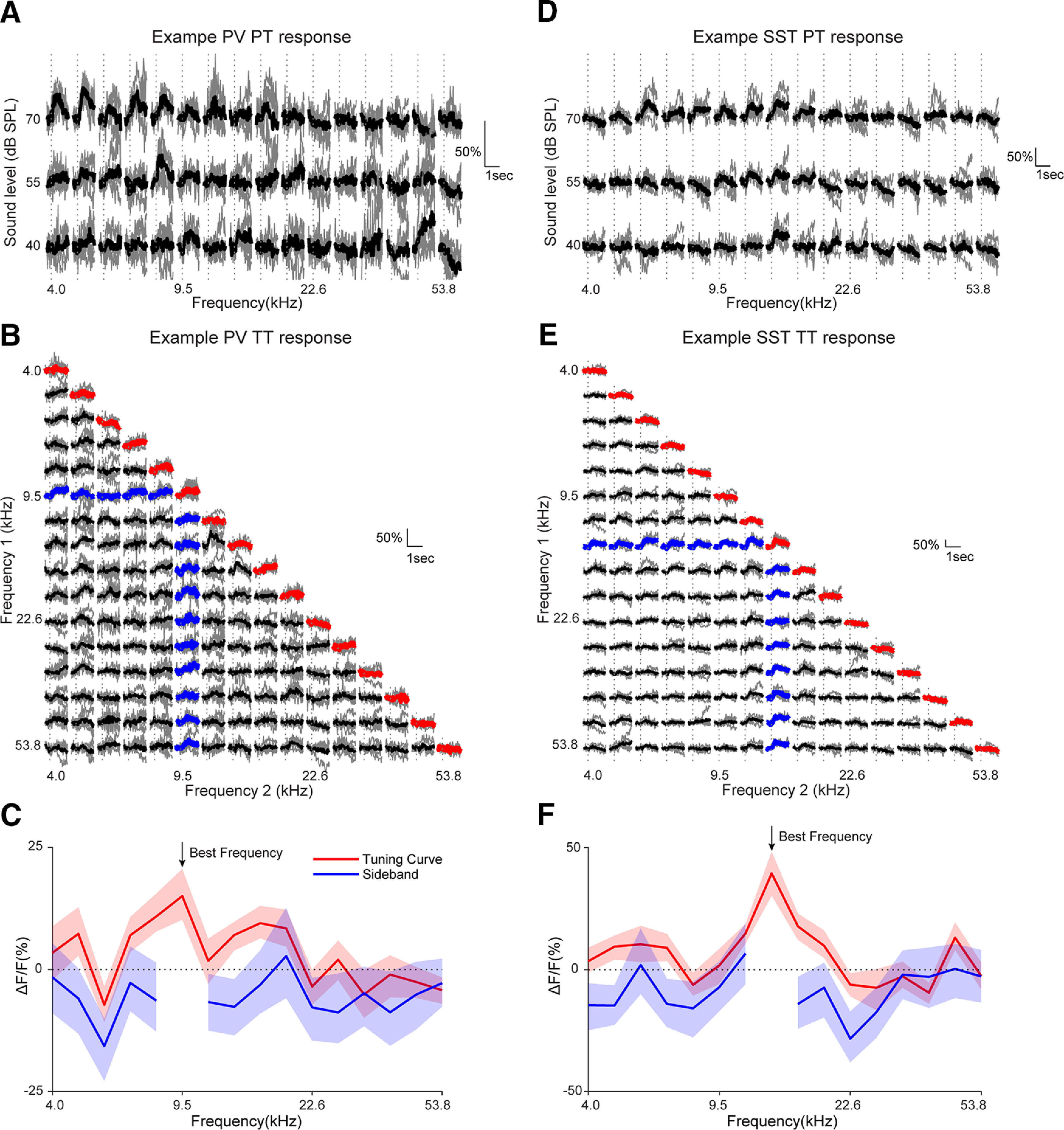
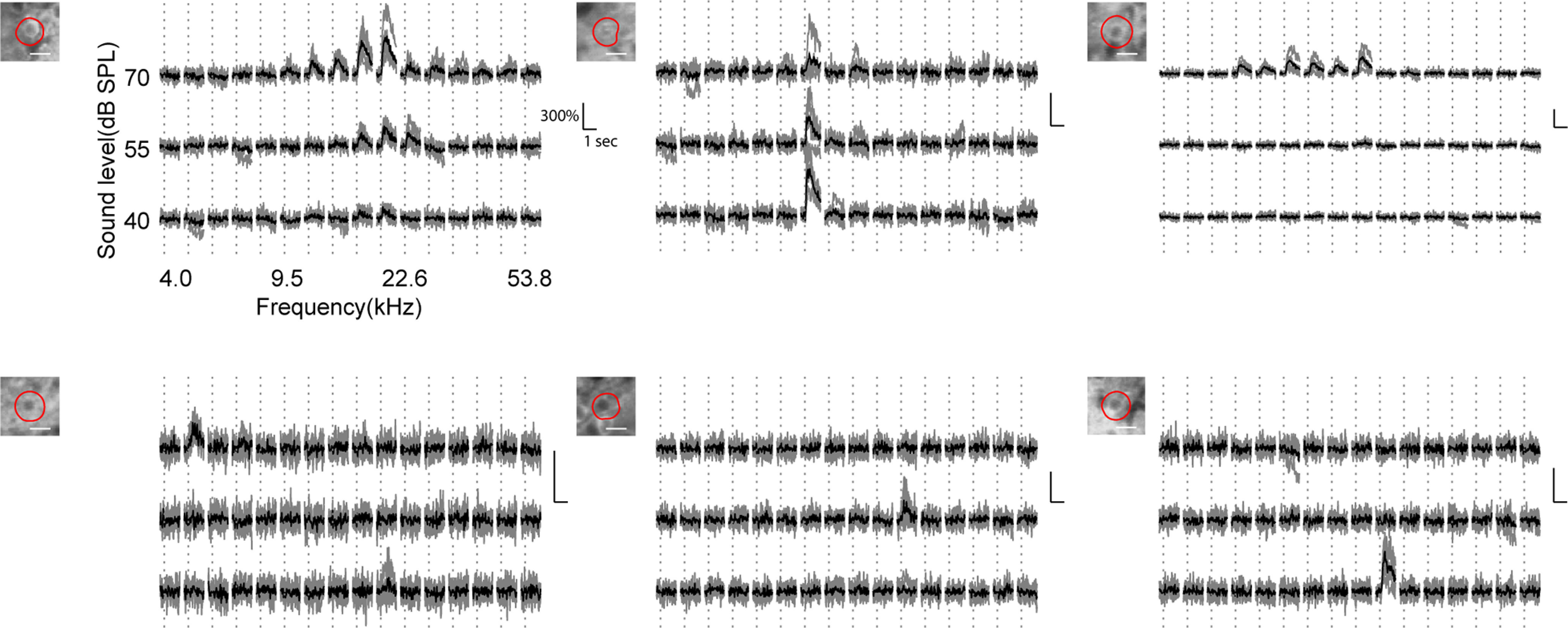
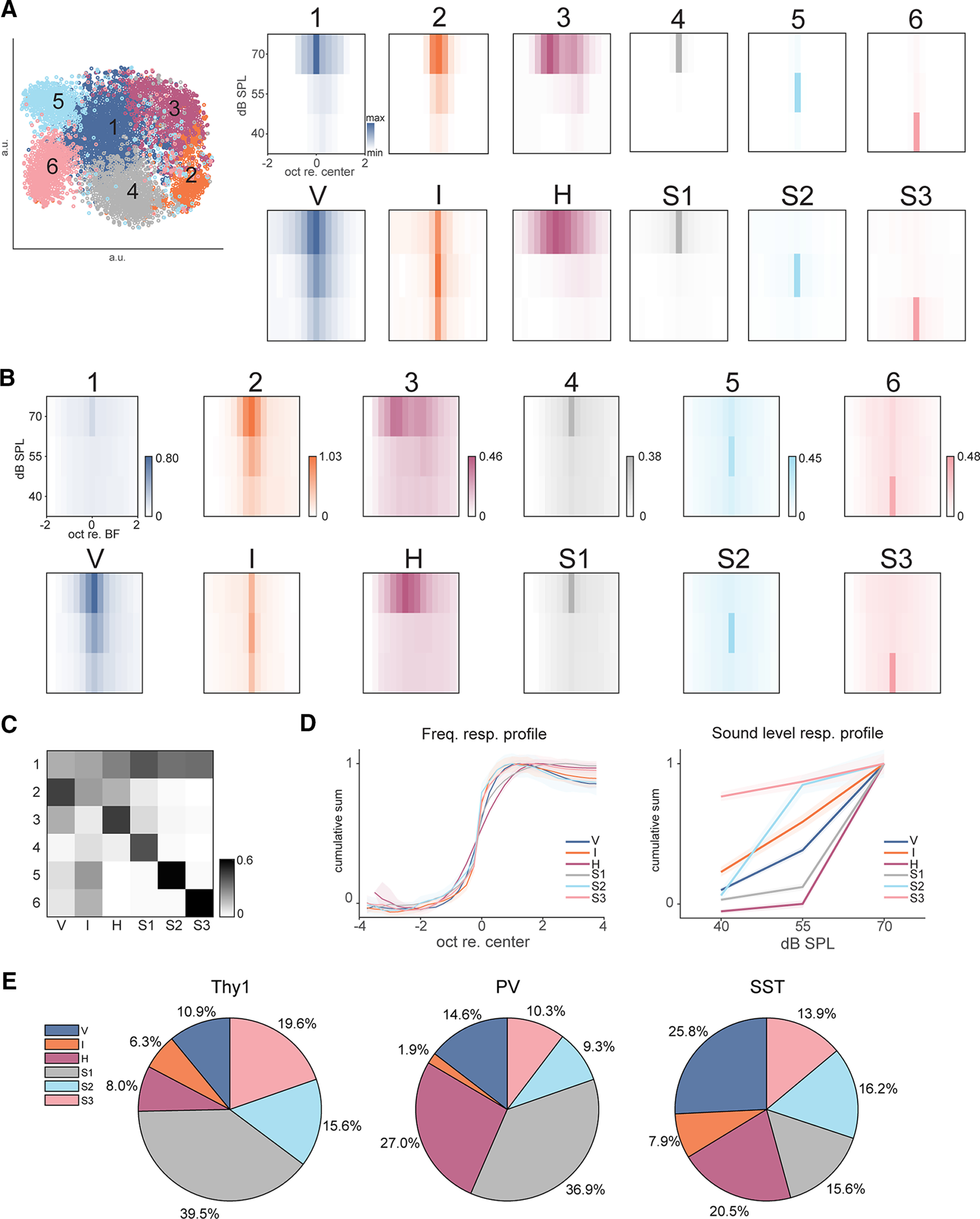
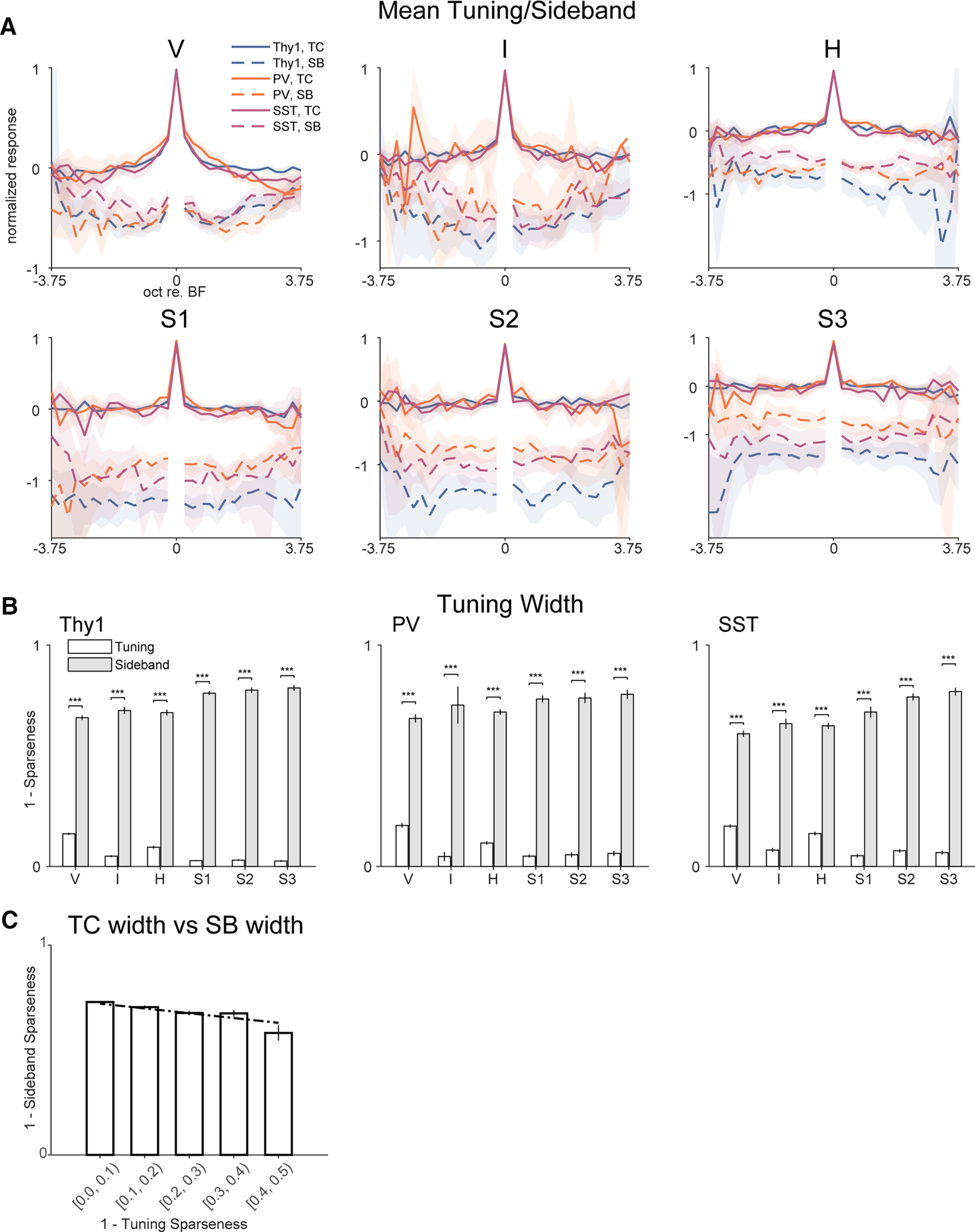
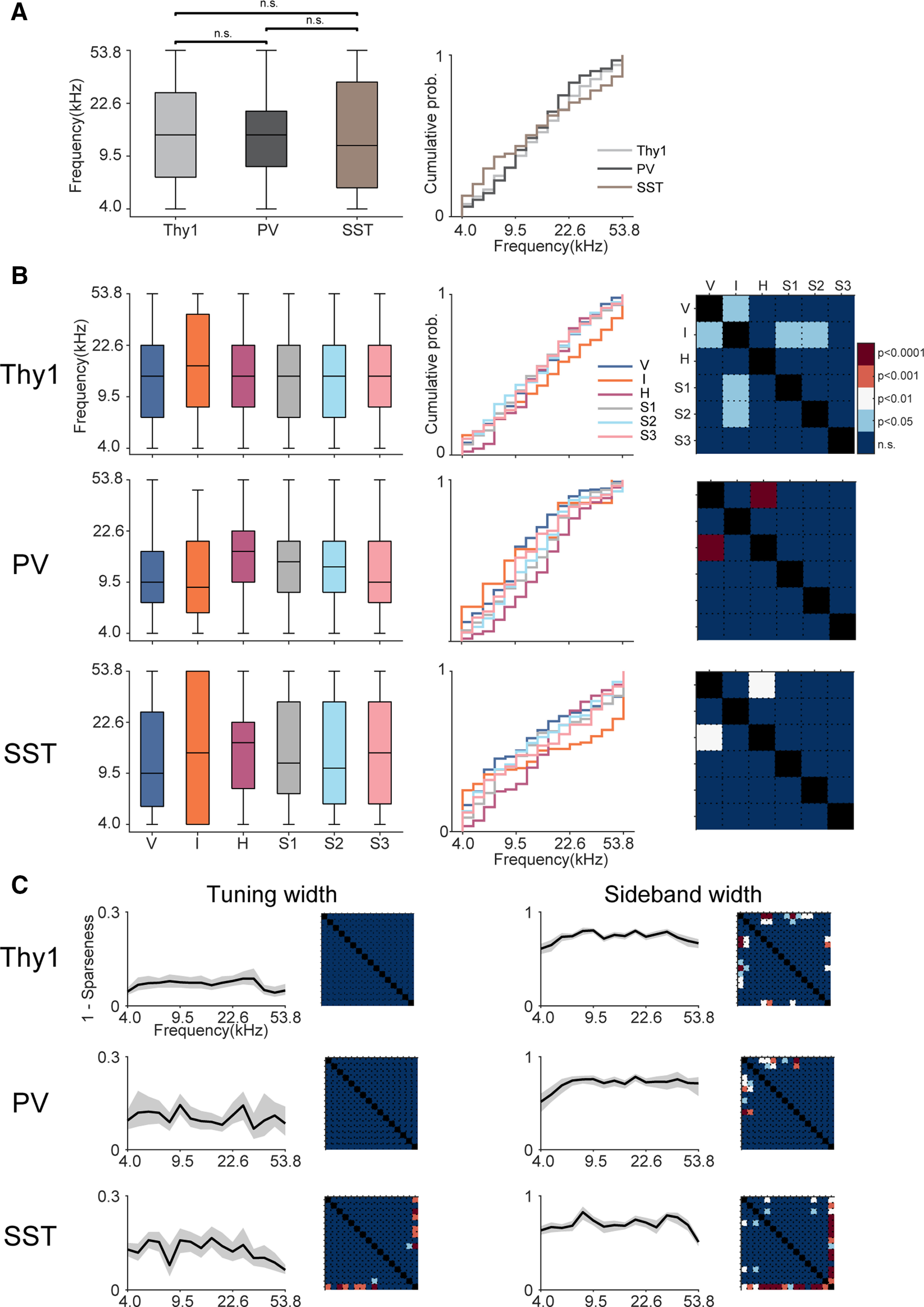
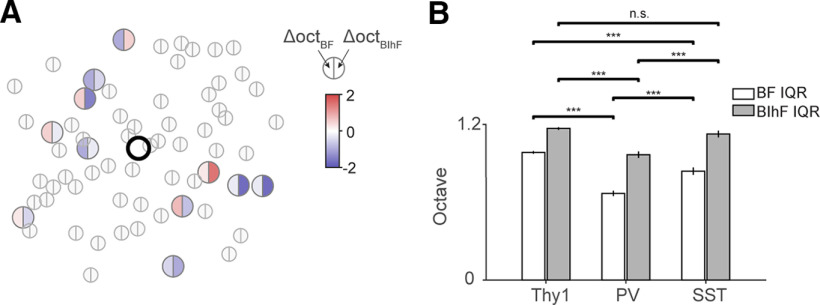
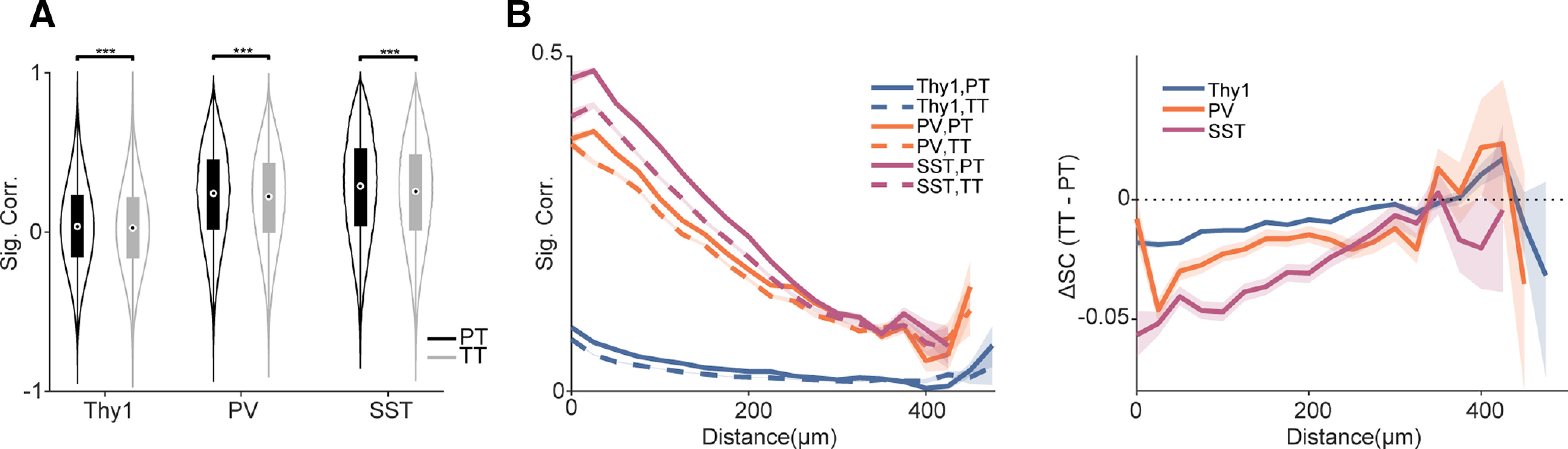
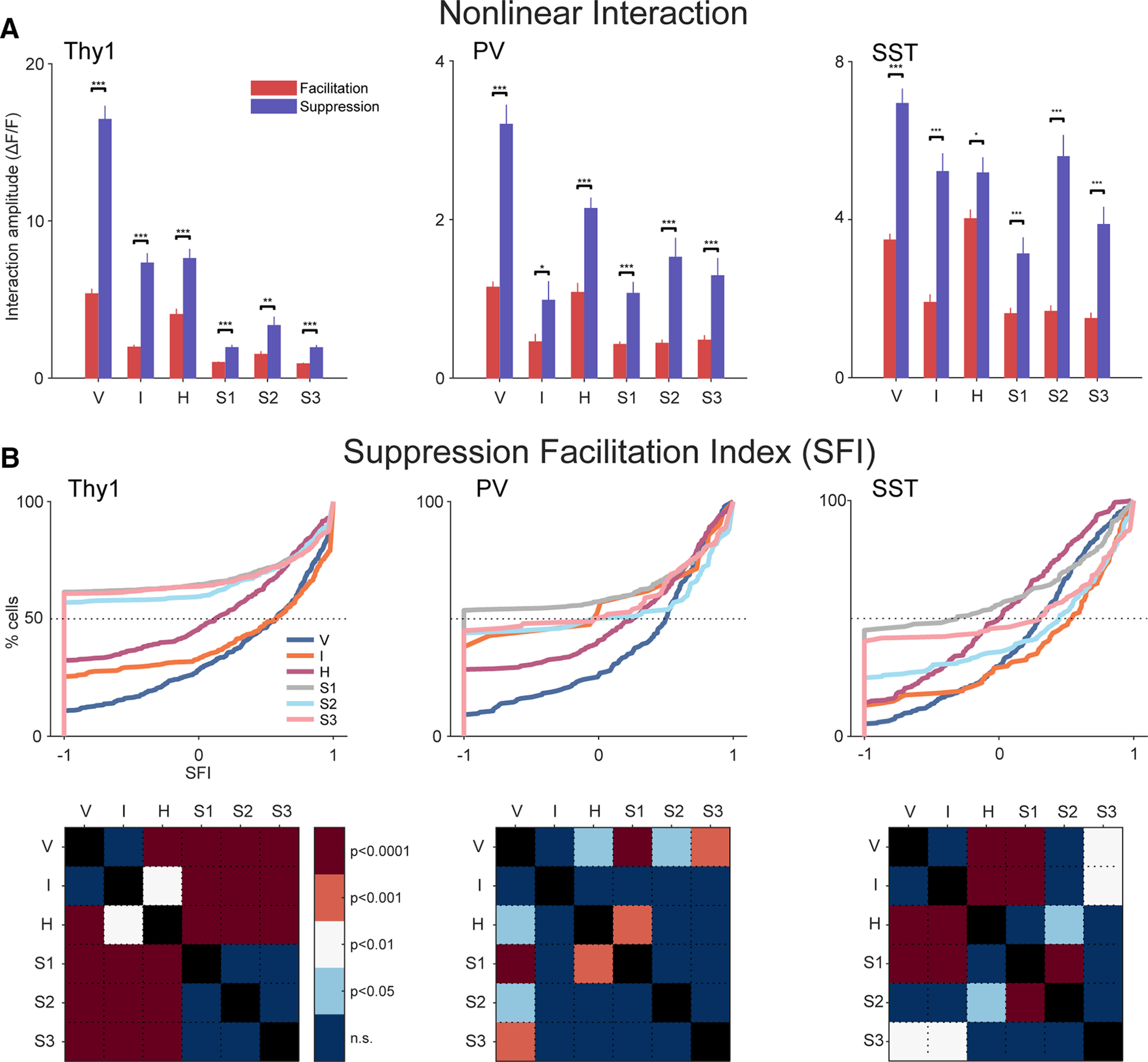
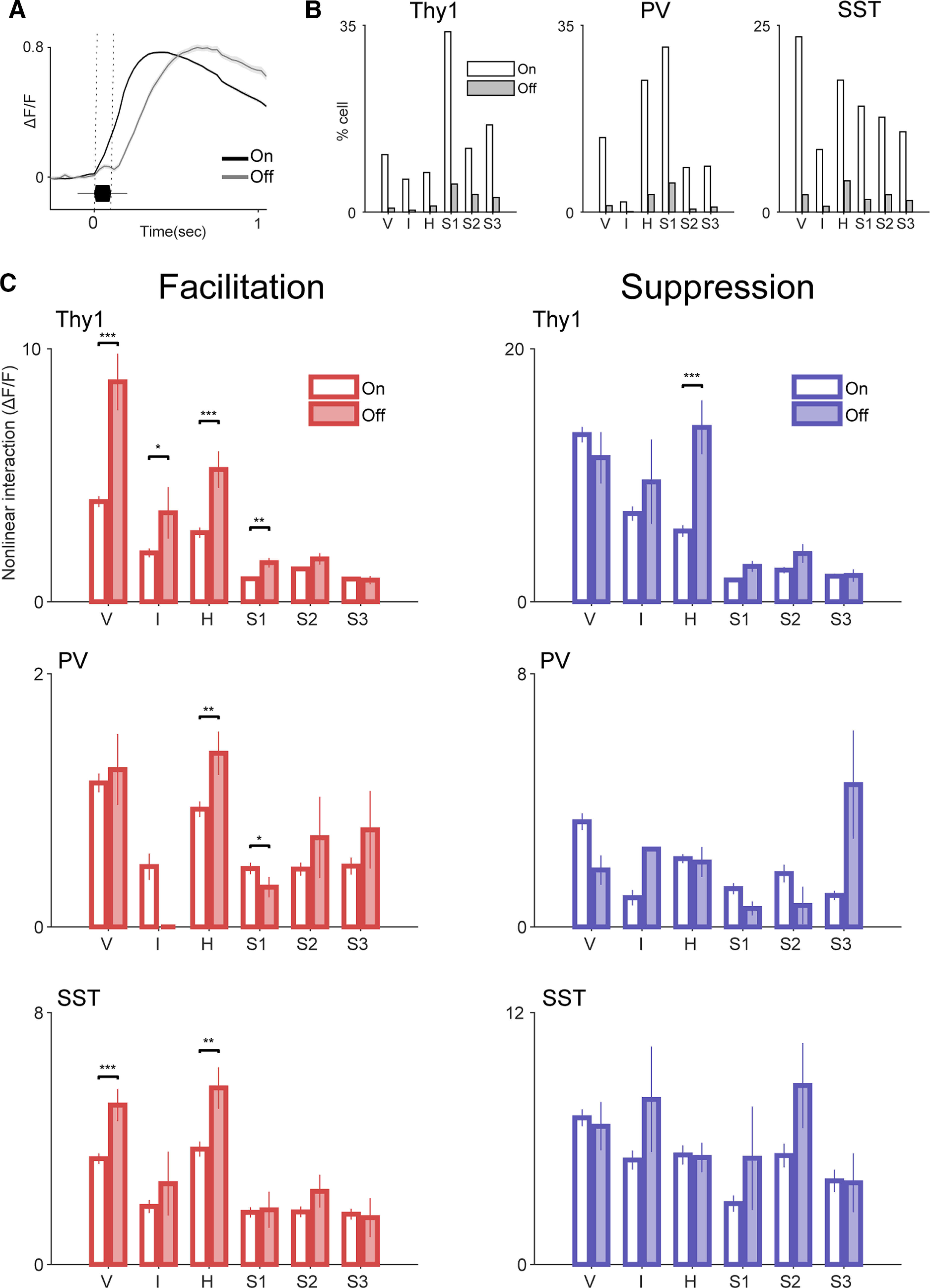
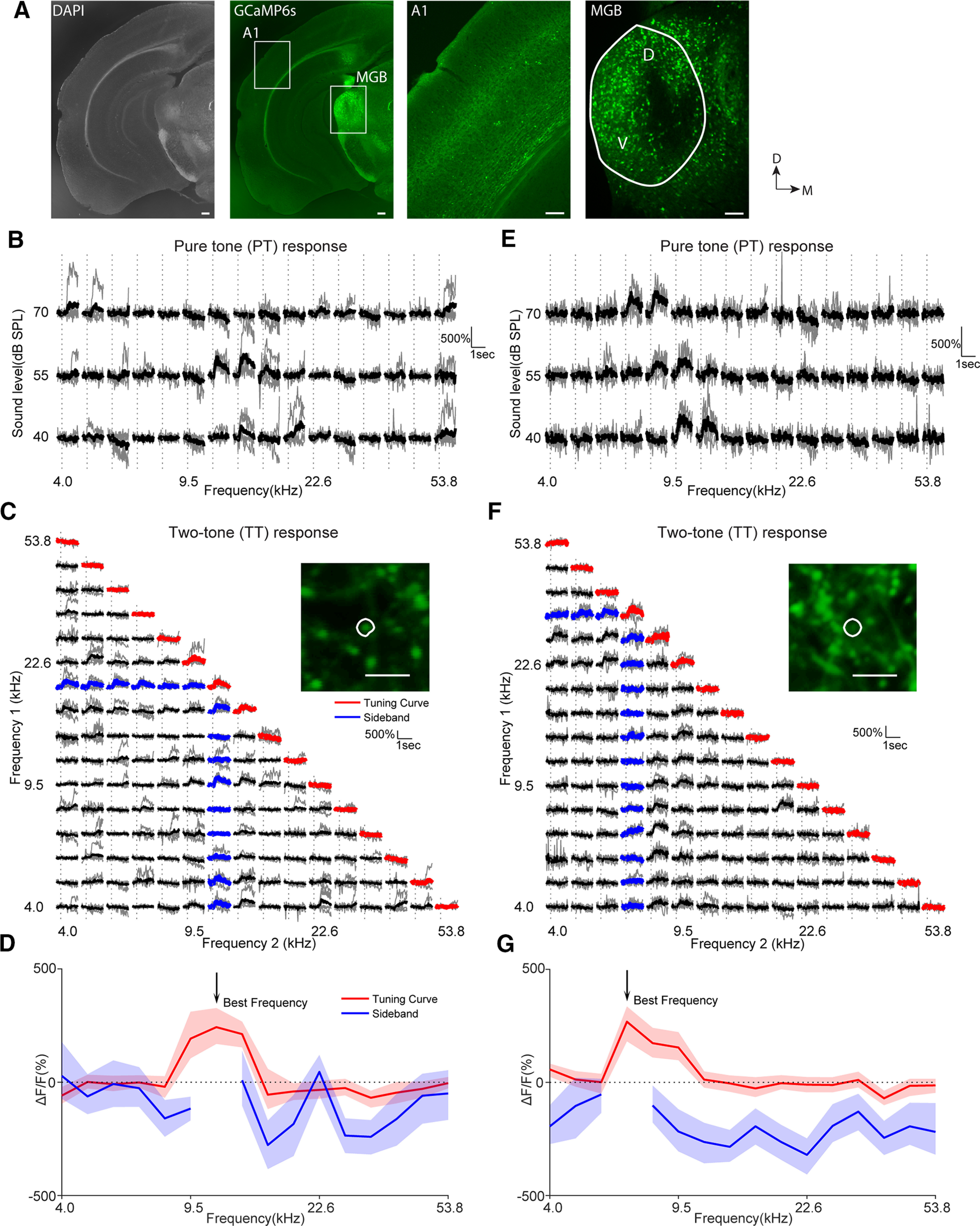
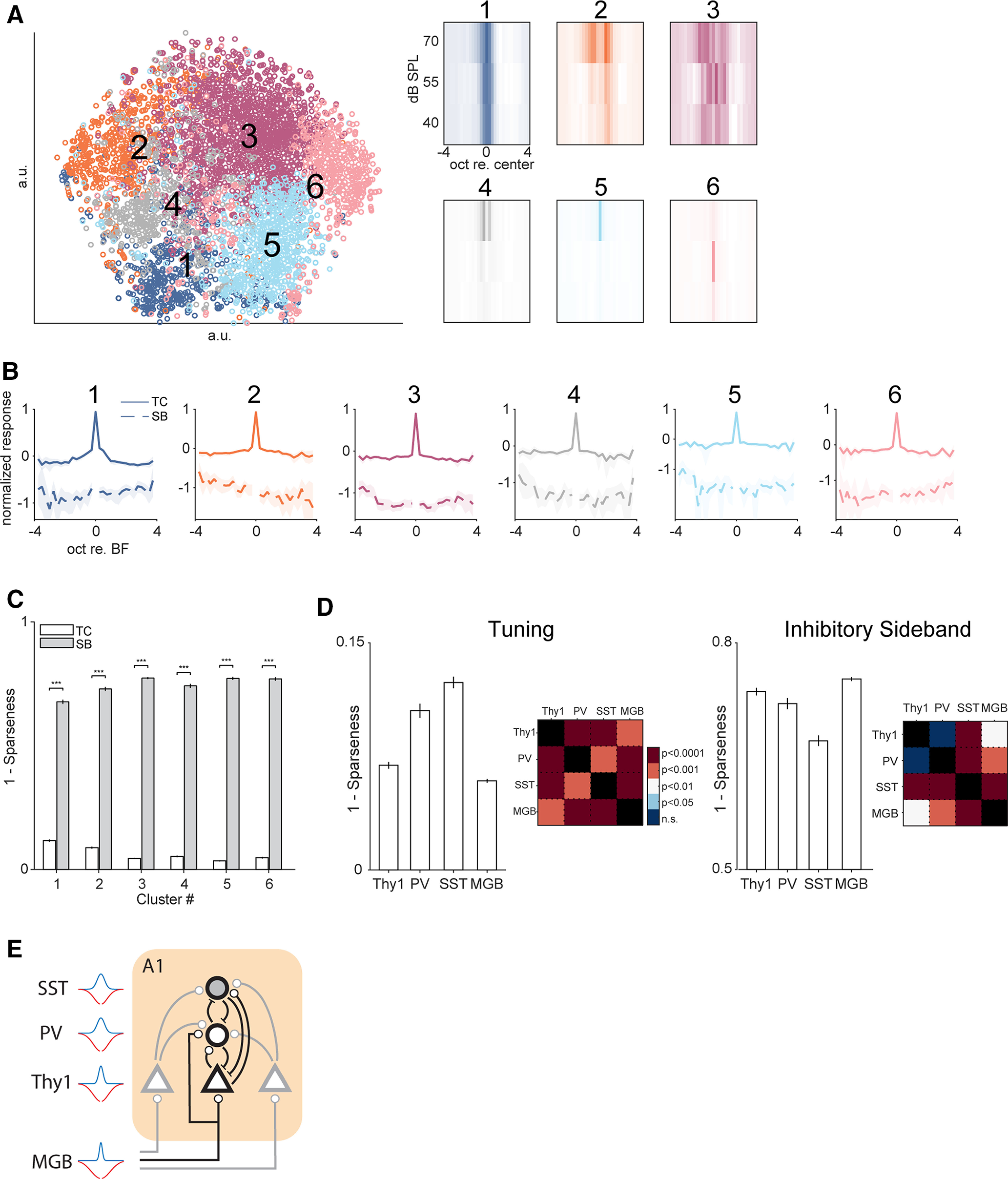
Receptive fields of primary auditory cortex (A1) neurons show excitatory neuronal frequency preference and diverse inhibitory sidebands. While the frequency preferences of excitatory neurons in local A1 areas can be heterogeneous, those of inhibitory neurons are more homogeneous. To date, the diversity and the origin of inhibitory sidebands in local neuronal populations and the relation between local cellular frequency preference and inhibitory sidebands are unknown. To reveal both excitatory and inhibitory subfields, we presented two-tone and pure tone stimuli while imaging excitatory neurons (Thy1) and two types of inhibitory neurons (parvalbumin and somatostatin) in L2/3 of mice A1. We classified neurons into six classes based on frequency response area (FRA) shapes and sideband inhibition depended both on FRA shapes and cell types. Sideband inhibition showed higher local heterogeneity than frequency tuning, suggesting that sideband inhibition originates from diverse sources of local and distant neurons. Two-tone interactions depended on neuron subclasses with excitatory neurons showing the most nonlinearity. Onset and offset neurons showed dissimilar spectral integration, suggesting differing circuits processing sound onset and offset. These results suggest that excitatory neurons integrate complex and nonuniform inhibitory input. Thalamocortical terminals also exhibited sideband inhibition, but with different properties from those of cortical neurons. Thus, some components of sideband inhibition are inherited from thalamocortical inputs and are further modified by converging intracortical circuits. The combined heterogeneity of frequency tuning and diverse sideband inhibition facilitates complex spectral shape encoding and allows for rapid and extensive plasticity.SIGNIFICANCE STATEMENT Sensory systems recognize and differentiate between different stimuli through selectivity for different features. Sideband inhibition serves as an important mechanism to sharpen stimulus selectivity, but its cortical mechanisms are not entirely resolved. We imaged pyramidal neurons and two common classes of interneurons suggested to mediate sideband inhibition (parvalbumin and somatostatin positive) in the auditory cortex and inferred their inhibitory sidebands. We observed a higher degree of variability in the inhibitory sideband than in the local frequency tuning, which cannot be predicted from the relative high homogeneity of responses by inhibitory interneurons. This suggests that cortical sideband inhibition is nonuniform and likely results from a complex interplay between existing functional inhibition in the feedforward input and cortical refinement.
Keywords: auditory cortex; inhibition; layer 2/3; mouse; sideband; tuning.
Copyright © 2021 the authors.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Differential Receptive Field Properties of Parvalbumin and Somatostatin Inhibitory Neurons in Mouse Auditory Cortex.Cereb Cortex. 2015 Jul;25(7):1782-91. doi: 10.1093/cercor/bht417. Epub 2014 Jan 14. Cereb Cortex. 2015. PMID: 24425250 Free PMC article.
-
Parvalbumin-expressing inhibitory interneurons in auditory cortex are well-tuned for frequency.J Neurosci. 2013 Aug 21;33(34):13713-23. doi: 10.1523/JNEUROSCI.0663-13.2013. J Neurosci. 2013. PMID: 23966693 Free PMC article.
-
Fine Control of Sound Frequency Tuning and Frequency Discrimination Acuity by Synaptic Zinc Signaling in Mouse Auditory Cortex.J Neurosci. 2019 Jan 30;39(5):854-865. doi: 10.1523/JNEUROSCI.1339-18.2018. Epub 2018 Nov 30. J Neurosci. 2019. PMID: 30504277 Free PMC article.
-
Development of auditory cortical synaptic receptive fields.Neurosci Biobehav Rev. 2011 Nov;35(10):2105-13. doi: 10.1016/j.neubiorev.2011.02.006. Epub 2011 Feb 15. Neurosci Biobehav Rev. 2011. PMID: 21329722 Free PMC article. Review.
-
Spectrotemporal dynamics of auditory cortical synaptic receptive field plasticity.Hear Res. 2011 Sep;279(1-2):149-61. doi: 10.1016/j.heares.2011.03.005. Epub 2011 Mar 21. Hear Res. 2011. PMID: 21426927 Free PMC article. Review.
Cited by
-
An inhibitory glycinergic projection from the cochlear nucleus to the lateral superior olive.Front Neural Circuits. 2023 Dec 1;17:1307283. doi: 10.3389/fncir.2023.1307283. eCollection 2023. Front Neural Circuits. 2023. PMID: 38107610 Free PMC article.
-
Perinatal Opioid Exposure Results in Persistent Hypoconnectivity of Excitatory Circuits and Reduced Activity Correlations in Mouse Primary Auditory Cortex.J Neurosci. 2022 Apr 27;42(17):3676-3687. doi: 10.1523/JNEUROSCI.2542-21.2022. Epub 2022 Mar 24. J Neurosci. 2022. PMID: 35332087 Free PMC article.
-
Auditory cortex ensembles jointly encode sound and locomotion speed to support sound perception during movement.PLoS Biol. 2023 Aug 31;21(8):e3002277. doi: 10.1371/journal.pbio.3002277. eCollection 2023 Aug. PLoS Biol. 2023. PMID: 37651461 Free PMC article.
-
Sparse representation of neurons for encoding complex sounds in the auditory cortex.Prog Neurobiol. 2024 Oct;241:102661. doi: 10.1016/j.pneurobio.2024.102661. Epub 2024 Sep 18. Prog Neurobiol. 2024. PMID: 39303758
-
Patchy harmonic functional connectivity of the mouse auditory cortex.Proc Natl Acad Sci U S A. 2025 Jul 8;122(27):e2510012122. doi: 10.1073/pnas.2510012122. Epub 2025 Jun 30. Proc Natl Acad Sci U S A. 2025. PMID: 40587804
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous