

Trichoderma reesei Rad51 tolerates mismatches in hybrid meiosis with diverse genome sequences
- PMID: 33593897
- PMCID: PMC7923544
- DOI: 10.1073/pnas.2007192118
Trichoderma reesei Rad51 tolerates mismatches in hybrid meiosis with diverse genome sequences
Abstract
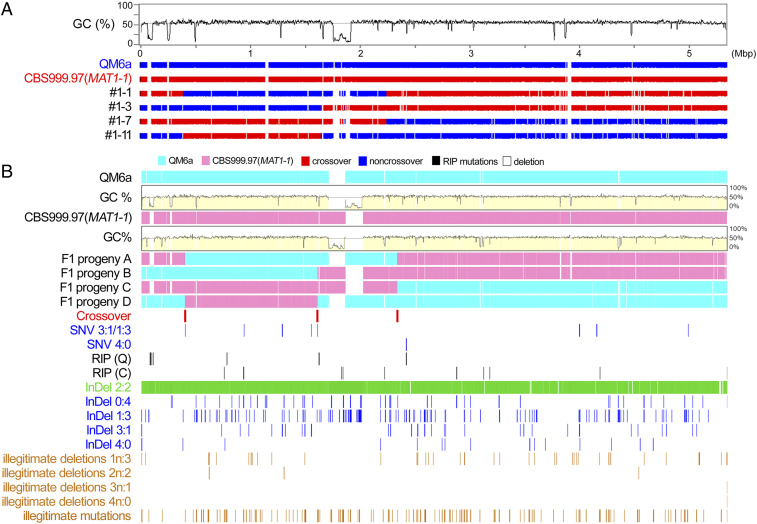
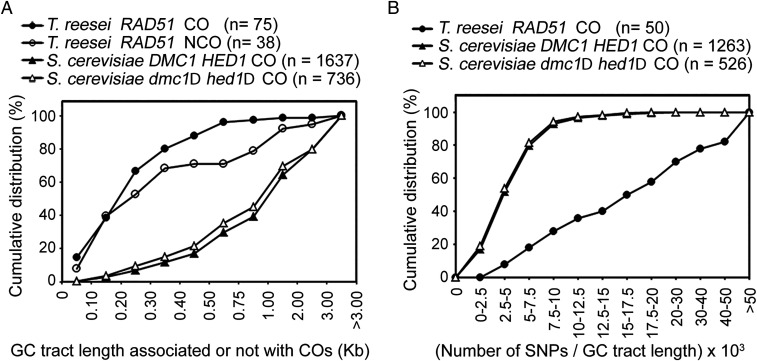
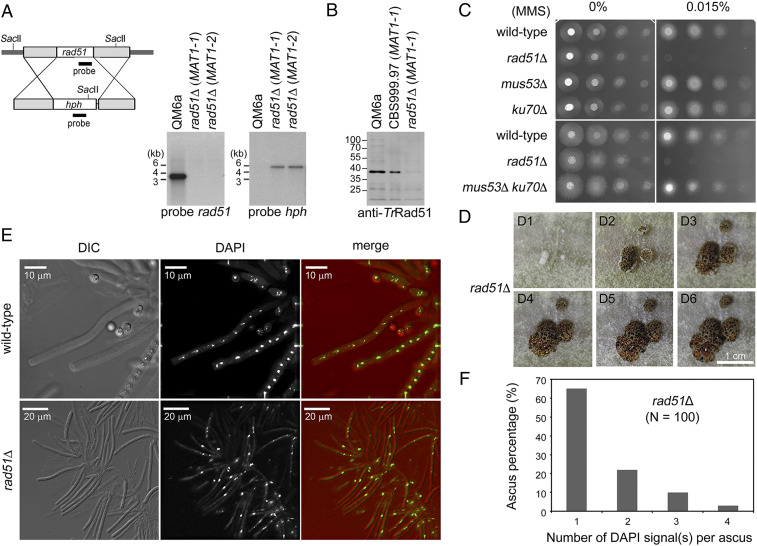
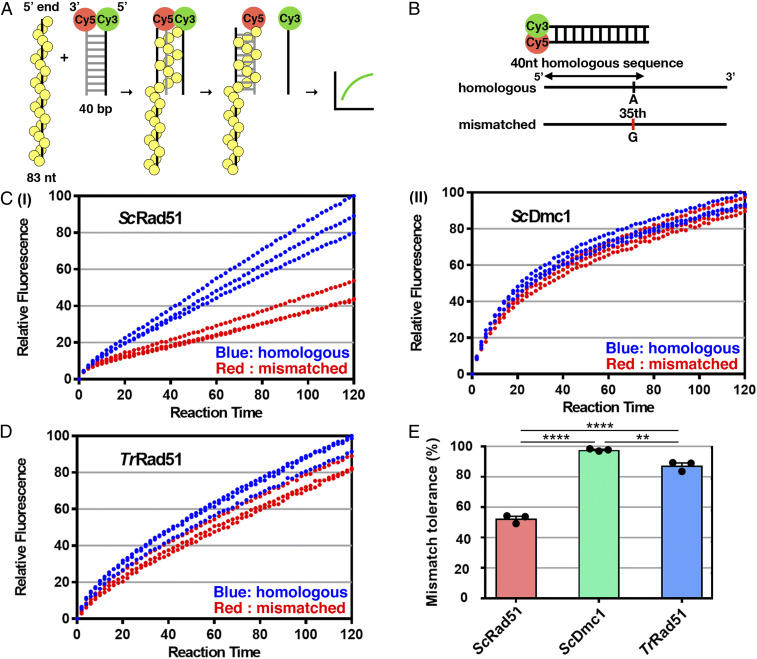
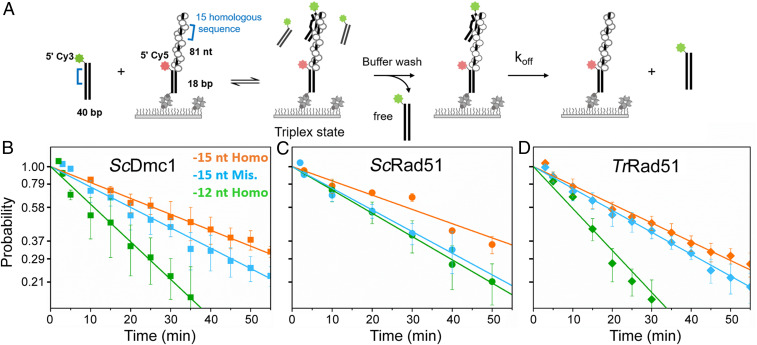
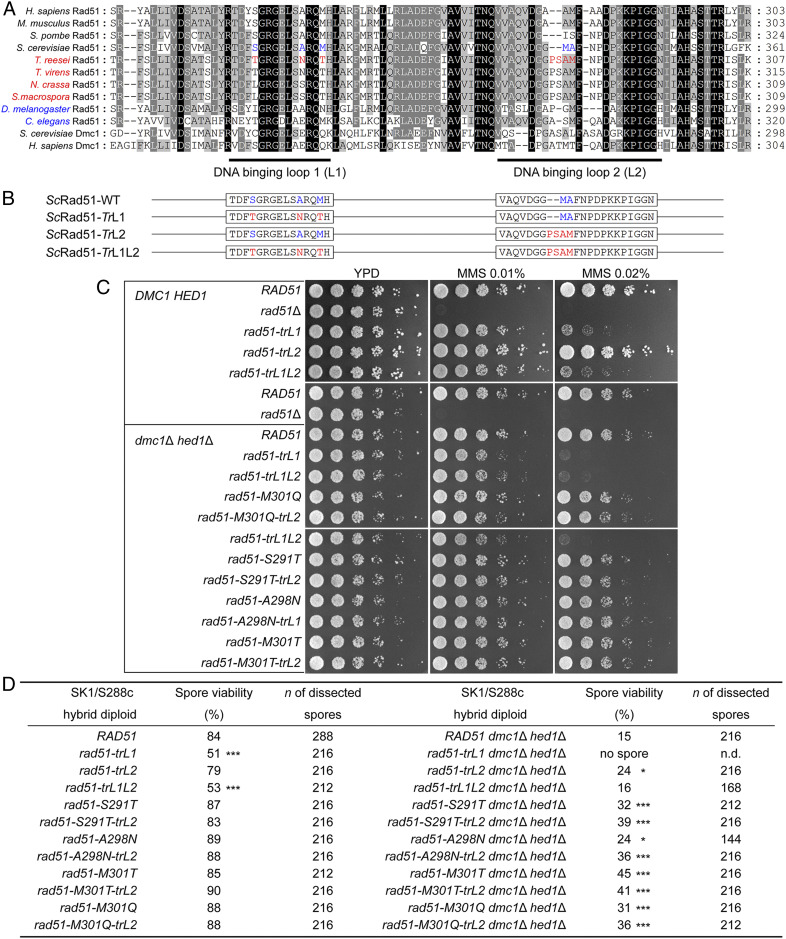
Most eukaryotes possess two RecA-like recombinases (ubiquitous Rad51 and meiosis-specific Dmc1) to promote interhomolog recombination during meiosis. However, some eukaryotes have lost Dmc1. Given that mammalian and yeast Saccharomyces cerevisiae (Sc) Dmc1 have been shown to stabilize recombination intermediates containing mismatches better than Rad51, we used the Pezizomycotina filamentous fungus Trichoderma reesei to address if and how Rad51-only eukaryotes conduct interhomolog recombination in zygotes with high sequence heterogeneity. We applied multidisciplinary approaches (next- and third-generation sequencing technology, genetics, cytology, bioinformatics, biochemistry, and single-molecule biophysics) to show that T. reesei Rad51 (TrRad51) is indispensable for interhomolog recombination during meiosis and, like ScDmc1, TrRad51 possesses better mismatch tolerance than ScRad51 during homologous recombination. Our results also indicate that the ancestral TrRad51 evolved to acquire ScDmc1-like properties by creating multiple structural variations, including via amino acid residues in the L1 and L2 DNA-binding loops.
Keywords: Dmc1; Rad51; Trichoderma; homologous recombination; meiosis.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
Rad51 facilitates filament assembly of meiosis-specific Dmc1 recombinase.Proc Natl Acad Sci U S A. 2020 May 26;117(21):11257-11264. doi: 10.1073/pnas.1920368117. Epub 2020 May 13. Proc Natl Acad Sci U S A. 2020. PMID: 32404423 Free PMC article.
-
The third exon of the budding yeast meiotic recombination gene HOP2 is required for calcium-dependent and recombinase Dmc1-specific stimulation of homologous strand assimilation.J Biol Chem. 2014 Jun 27;289(26):18076-86. doi: 10.1074/jbc.M114.558601. Epub 2014 May 5. J Biol Chem. 2014. PMID: 24798326 Free PMC article.
-
Spontaneous self-segregation of Rad51 and Dmc1 DNA recombinases within mixed recombinase filaments.J Biol Chem. 2018 Mar 16;293(11):4191-4200. doi: 10.1074/jbc.RA117.001143. Epub 2018 Jan 30. J Biol Chem. 2018. PMID: 29382724 Free PMC article.
-
From meiosis to postmeiotic events: uncovering the molecular roles of the meiosis-specific recombinase Dmc1.FEBS J. 2010 Feb;277(3):590-8. doi: 10.1111/j.1742-4658.2009.07503.x. Epub 2009 Dec 15. FEBS J. 2010. PMID: 20015079 Review.
-
The biochemistry of early meiotic recombination intermediates.Cell Cycle. 2018;17(23):2520-2530. doi: 10.1080/15384101.2018.1553355. Epub 2018 Dec 10. Cell Cycle. 2018. PMID: 30482074 Free PMC article. Review.
Cited by
-
Trichoderma - genomes and genomics as treasure troves for research towards biology, biotechnology and agriculture.Front Fungal Biol. 2022 Sep 14;3:1002161. doi: 10.3389/ffunb.2022.1002161. eCollection 2022. Front Fungal Biol. 2022. PMID: 37746224 Free PMC article. Review.
-
Mei5-Sae3 stabilizes Dmc1 nucleating clusters for efficient Dmc1 assembly on RPA-coated single-stranded DNA.Nucleic Acids Res. 2024 Oct 28;52(19):11768-11784. doi: 10.1093/nar/gkae780. Nucleic Acids Res. 2024. PMID: 39275989 Free PMC article.
-
Hop2-Mnd1 functions as a DNA sequence fidelity switch in Dmc1-mediated DNA recombination.Nat Commun. 2024 Oct 27;15(1):9266. doi: 10.1038/s41467-024-53641-3. Nat Commun. 2024. PMID: 39463417 Free PMC article.
-
Mechanisms of distinctive mismatch tolerance between Rad51 and Dmc1 in homologous recombination.Nucleic Acids Res. 2021 Dec 16;49(22):13135-13149. doi: 10.1093/nar/gkab1141. Nucleic Acids Res. 2021. PMID: 34871438 Free PMC article.
-
Trichoderma species from plant and soil: An excellent resource for biosynthesis of terpenoids with versatile bioactivities.J Adv Res. 2023 Jul;49:81-102. doi: 10.1016/j.jare.2022.09.010. Epub 2022 Oct 1. J Adv Res. 2023. PMID: 36195283 Free PMC article. Review.
References
-
- Allers T., Lichten M., Differential timing and control of noncrossover and crossover recombination during meiosis. Cell 106, 47–57 (2001). - PubMed
-
- Börner G. V., Kleckner N., Hunter N., Crossover/noncrossover differentiation, synaptonemal complex formation, and regulatory surveillance at the leptotene/zygotene transition of meiosis. Cell 117, 29–45 (2004). - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
