

Type I Interferon Signaling Is a Common Factor Driving Streptococcus pneumoniae and Influenza A Virus Shedding and Transmission
- PMID: 33593970
- PMCID: PMC8545127
- DOI: 10.1128/mBio.03589-20
Type I Interferon Signaling Is a Common Factor Driving Streptococcus pneumoniae and Influenza A Virus Shedding and Transmission
Abstract
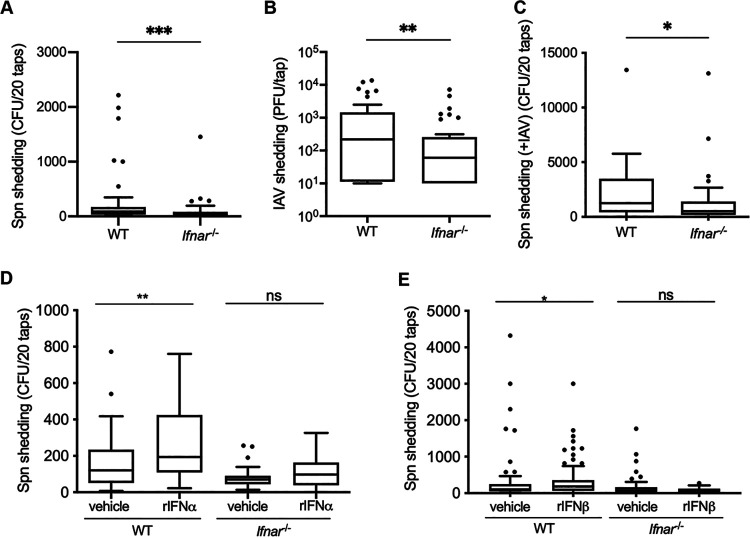
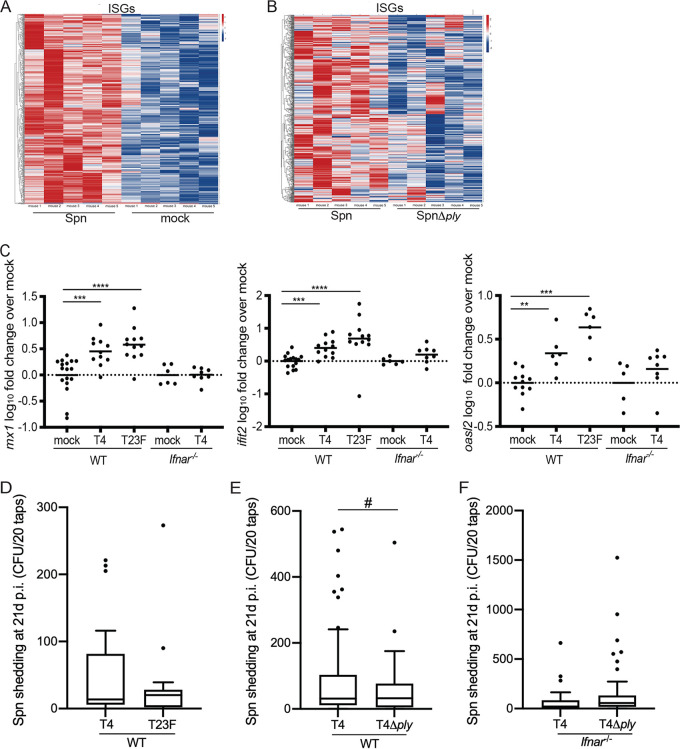
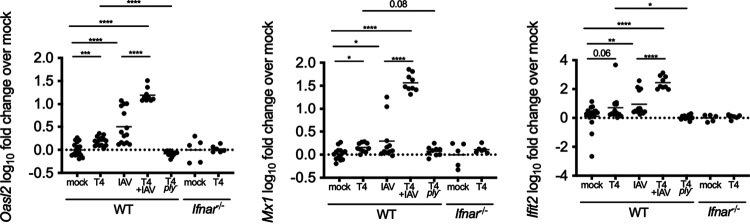
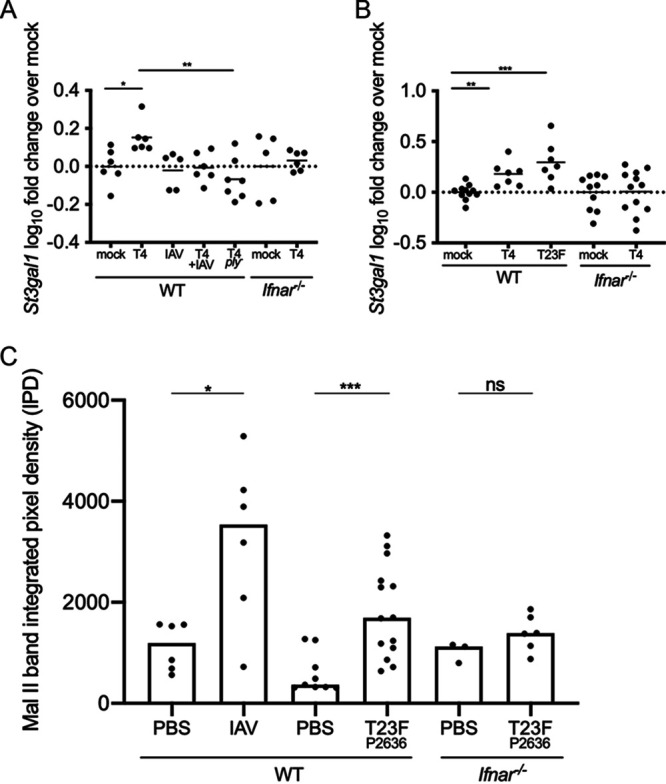
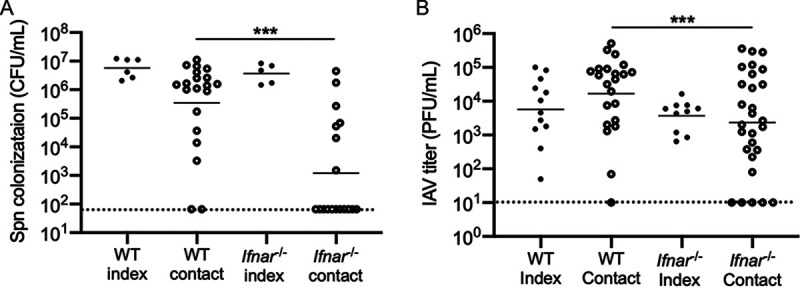
The dynamics underlying respiratory contagion (the transmission of infectious agents from the airways) are poorly understood. We investigated host factors involved in the transmission of the leading respiratory pathogen Streptococcus pneumoniae Using an infant mouse model, we examined whether S. pneumoniae triggers inflammatory pathways shared by influenza A virus (IAV) to promote nasal secretions and shedding from the upper respiratory tract to facilitate transit to new hosts. Here, we show that amplification of the type I interferon (IFN-I) response is a critical host factor in this process, as shedding and transmission by both IAV and S. pneumoniae were decreased in pups lacking the common IFN-I receptor (Ifnar1-/- mice). Additionally, providing exogenous recombinant IFN-I to S. pneumoniae-infected pups was sufficient to increase bacterial shedding. The expression of IFN-stimulated genes (ISGs) was upregulated in S. pneumoniae-infected wild-type (WT) but not Ifnar1-/- mice, including genes involved in mucin type O-glycan biosynthesis; this correlated with an increase in secretions in S. pneumoniae- and IAV-infected WT compared to Ifnar1-/- pups. S. pneumoniae stimulation of ISGs was largely dependent on its pore-forming toxin, pneumolysin, and coinfection with IAV and S. pneumoniae resulted in synergistic increases in ISG expression. We conclude that the induction of IFN-I signaling appears to be a common factor driving viral and bacterial respiratory contagion.IMPORTANCE Respiratory tract infections are a leading cause of childhood mortality and, globally, Streptococcus pneumoniae is the leading cause of mortality due to pneumonia. Transmission of S. pneumoniae primarily occurs through direct contact with respiratory secretions, although the host and bacterial factors underlying transmission are poorly understood. We examined transmission dynamics of S. pneumoniae in an infant mouse model and here show that S. pneumoniae colonization of the upper respiratory tract stimulates host inflammatory pathways commonly associated with viral infections. Amplification of this response was shown to be a critical host factor driving shedding and transmission of both S. pneumoniae and influenza A virus, with infection stimulating expression of a wide variety of genes, including those involved in the biosynthesis of mucin, a major component of respiratory secretions. Our findings suggest a mechanism facilitating S. pneumoniae contagion that is shared by viral infection.
Keywords: influenza virus; interferon; pneumococcus; transmission.
Copyright © 2021 Zangari et al.
Figures
Similar articles
-
An Infant Mouse Model of Influenza Virus Transmission Demonstrates the Role of Virus-Specific Shedding, Humoral Immunity, and Sialidase Expression by Colonizing Streptococcus pneumoniae.mBio. 2018 Dec 18;9(6):e02359-18. doi: 10.1128/mBio.02359-18. mBio. 2018. PMID: 30563897 Free PMC article.
-
TLR2 signaling decreases transmission of Streptococcus pneumoniae by limiting bacterial shedding in an infant mouse Influenza A co-infection model.PLoS Pathog. 2014 Aug 28;10(8):e1004339. doi: 10.1371/journal.ppat.1004339. eCollection 2014 Aug. PLoS Pathog. 2014. PMID: 25166617 Free PMC article.
-
Synergism and Antagonism of Bacterial-Viral Coinfection in the Upper Respiratory Tract.mSphere. 2022 Feb 23;7(1):e0098421. doi: 10.1128/msphere.00984-21. Epub 2022 Jan 19. mSphere. 2022. PMID: 35044807 Free PMC article.
-
Inflammation as a Modulator of Host Susceptibility to Pulmonary Influenza, Pneumococcal, and Co-Infections.Front Immunol. 2020 Feb 11;11:105. doi: 10.3389/fimmu.2020.00105. eCollection 2020. Front Immunol. 2020. PMID: 32117259 Free PMC article. Review.
-
Altered Signal Transduction in the Immune Response to Influenza Virus and S. pneumoniae or S. aureus Co-Infections.Int J Mol Sci. 2021 May 22;22(11):5486. doi: 10.3390/ijms22115486. Int J Mol Sci. 2021. PMID: 34067487 Free PMC article. Review.
Cited by
-
Bacterial surface lipoproteins mediate epithelial microinvasion by Streptococcus pneumoniae.Infect Immun. 2024 May 7;92(5):e0044723. doi: 10.1128/iai.00447-23. Epub 2024 Apr 17. Infect Immun. 2024. PMID: 38629841 Free PMC article.
-
Streptococcus pneumoniae colonization associates with impaired adaptive immune responses against SARS-CoV-2.J Clin Invest. 2022 Apr 1;132(7):e157124. doi: 10.1172/JCI157124. J Clin Invest. 2022. PMID: 35139037 Free PMC article.
-
Type I IFN Signaling Is Essential for Preventing IFN-γ Hyperproduction and Subsequent Deterioration of Antibacterial Immunity during Postinfluenza Pneumococcal Infection.J Immunol. 2022 Jul 1;209(1):128-135. doi: 10.4049/jimmunol.2101135. Epub 2022 Jun 15. J Immunol. 2022. PMID: 35705254 Free PMC article.
-
A bacterial vesicle-based pneumococcal vaccine against influenza-mediated secondary Streptococcus pneumoniae pulmonary infection.Mucosal Immunol. 2024 Apr;17(2):169-181. doi: 10.1016/j.mucimm.2024.01.002. Epub 2024 Jan 11. Mucosal Immunol. 2024. PMID: 38215909 Free PMC article.
-
The Two-Component System YesMN Promotes Pneumococcal Host-to-Host Transmission and Regulates Genes Involved in Zinc Homeostasis.Infect Immun. 2023 Jan 24;91(1):e0037522. doi: 10.1128/iai.00375-22. Epub 2022 Dec 20. Infect Immun. 2023. PMID: 36537790 Free PMC article.
References
-
- Ortigoza MB, Blaser SB, Zafar MA, Hammond AJ, Weiser JN. 2018. An infant mouse model of influenza virus transmission demonstrates the role of virus-specific shedding, humoral immunity, and sialidase expression by colonizing Streptococcus pneumoniae. mBio 9:e02359-18. doi:10.1128/mBio.02359-18. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
