

IL-27 Negatively Regulates Tip-DC Development during Infection
- PMID: 33593983
- PMCID: PMC8545113
- DOI: 10.1128/mBio.03385-20
IL-27 Negatively Regulates Tip-DC Development during Infection
Abstract
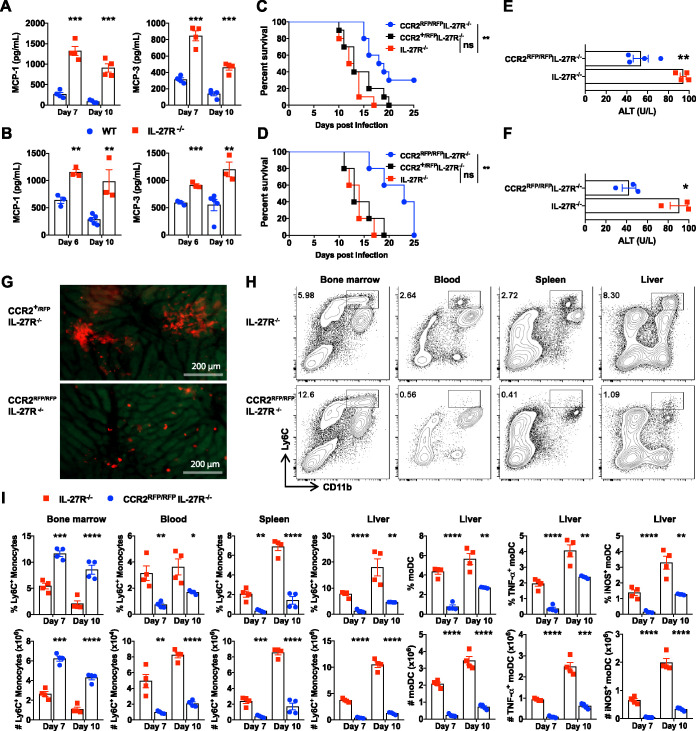
Tumor necrosis factor (TNF)/inducible nitric oxide synthase (iNOS)-producing dendritic cells (Tip-DCs) have profound impacts on host immune responses during infections. The mechanisms regulating Tip-DC development remain largely unknown. Here, using a mouse model of infection with African trypanosomes, we show that a deficiency in interleukin-27 receptor (IL-27R) signaling results in escalated intrahepatic accumulation of Ly6C-positive (Ly6C+) monocytes and their differentiation into Tip-DCs. Blocking Tip-DC development significantly ameliorates liver injury and increases the survival of infected IL-27R-/- mice. Mechanistically, Ly6C+ monocyte differentiation into pathogenic Tip-DCs in infected IL-27R-/- mice is driven by a CD4+ T cell-interferon gamma (IFN-γ) axis via cell-intrinsic IFN-γ signaling. In parallel, hyperactive IFN-γ signaling induces cell death of Ly6C-negative (Ly6C-) monocytes in a cell-intrinsic manner, which in turn aggravates the development of pathogenic Tip-DCs due to the loss of the negative regulation of Ly6C- monocytes on Ly6C+ monocyte differentiation into Tip-DCs. Thus, IL-27 inhibits the dual-track exacerbation of Tip-DC development induced by a CD4+ T cell-IFN-γ axis. We conclude that IL-27 negatively regulates Tip-DC development by preventing the cell-intrinsic effects of IFN-γ and that the regulation involves CD4+ T cells and Ly6C- monocytes. Targeting IL-27 signaling may manipulate Tip-DC development for therapeutic intervention.IMPORTANCE TNF/iNOS-producing dendritic cells (Tip-DCs) are at the front line as immune effector cells to fight off a broad range of invading microbes. Excessive development of Tip-DCs contributes to tissue destruction. Thus, identifying master regulators of Tip-DC development is fundamental for developing new therapeutic strategies. Here, we identify Tip-DCs as a terminal target of IL-27, which prevents Tip-DC-mediated early mortality during parasitic infections. We demonstrate that IL-27 inhibits Tip-DC development via a dual-track mechanism involving the complex interactions of effector CD4+ T cells, Ly6C- monocytes, and Ly6C+ monocytes. These findings delineate an in-depth view of mechanisms of Tip-DC differentiation that may have significant implications for the ongoing development of IL-27-based immunotherapy.
Keywords: African trypanosomes; IFN-γ; IL-27; Ly6C+ monocytes; Ly6C− monocytes; Tip-DCs; host-pathogen interactions; intravital imaging; liver immunity; murine model of African trypanosomiasis; parasites.
Copyright © 2021 Liu et al.
Figures





Similar articles
-
Tip-DC development during parasitic infection is regulated by IL-10 and requires CCL2/CCR2, IFN-gamma and MyD88 signaling.PLoS Pathog. 2010 Aug 12;6(8):e1001045. doi: 10.1371/journal.ppat.1001045. PLoS Pathog. 2010. PMID: 20714353 Free PMC article.
-
IL-10 dampens TNF/inducible nitric oxide synthase-producing dendritic cell-mediated pathogenicity during parasitic infection.J Immunol. 2009 Jan 15;182(2):1107-18. doi: 10.4049/jimmunol.182.2.1107. J Immunol. 2009. PMID: 19124754
-
Human CD8⁺ T cells drive Th1 responses through the differentiation of TNF/iNOS-producing dendritic cells.Eur J Immunol. 2011 Jun;41(6):1639-51. doi: 10.1002/eji.201041022. Epub 2011 May 9. Eur J Immunol. 2011. PMID: 21469104
-
Differentiation and function of mouse monocyte-derived dendritic cells in steady state and inflammation.Immunol Rev. 2010 Mar;234(1):90-104. doi: 10.1111/j.0105-2896.2009.00876.x. Immunol Rev. 2010. PMID: 20193014 Review.
-
Role of cytokines in Trypanosoma brucei-induced anaemia: A review of the literature.Malawi Med J. 2015 Jun;27(2):45-50. doi: 10.4314/mmj.v27i2.3. Malawi Med J. 2015. PMID: 26405511 Free PMC article. Review.
Cited by
-
The Role of MIF and IL-10 as Molecular Yin-Yang in the Modulation of the Host Immune Microenvironment During Infections: African Trypanosome Infections as a Paradigm.Front Immunol. 2022 Apr 7;13:865395. doi: 10.3389/fimmu.2022.865395. eCollection 2022. Front Immunol. 2022. PMID: 35464430 Free PMC article. Review.
-
CXCR6+CD4+ T cells promote mortality during Trypanosoma brucei infection.PLoS Pathog. 2021 Oct 6;17(10):e1009968. doi: 10.1371/journal.ppat.1009968. eCollection 2021 Oct. PLoS Pathog. 2021. PMID: 34614031 Free PMC article.
-
IL-27 limits HSPC differentiation during infection and protects from stem cell exhaustion.bioRxiv [Preprint]. 2025 Jun 24:2025.01.15.633135. doi: 10.1101/2025.01.15.633135. bioRxiv. 2025. PMID: 39868131 Free PMC article. Preprint.
-
EGR3-HDAC6-IL-27 Axis Mediates Allergic Inflammation and Is Necessary for Tumorigenic Potential of Cancer Cells Enhanced by Allergic Inflammation-Promoted Cellular Interactions.Front Immunol. 2021 Jun 21;12:680441. doi: 10.3389/fimmu.2021.680441. eCollection 2021. Front Immunol. 2021. PMID: 34234781 Free PMC article.
-
An updated advancement of bifunctional IL-27 in inflammatory autoimmune diseases.Front Immunol. 2024 Mar 19;15:1366377. doi: 10.3389/fimmu.2024.1366377. eCollection 2024. Front Immunol. 2024. PMID: 38566992 Free PMC article. Review.
References
-
- Ziegler-Heitbrock L, Ancuta P, Crowe S, Dalod M, Grau V, Hart DN, Leenen PJM, Liu Y-J, MacPherson G, Randolph GJ, Scherberich J, Schmitz J, Shortman K, Sozzani S, Strobl H, Zembala M, Austyn JM, Lutz MB. 2010. Nomenclature of monocytes and dendritic cells in blood. Blood 116:e74–e80. doi:10.1182/blood-2010-02-258558. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
