

ABCC6 deficiency promotes dyslipidemia and atherosclerosis
- PMID: 33594095
- PMCID: PMC7887252
- DOI: 10.1038/s41598-021-82966-y
ABCC6 deficiency promotes dyslipidemia and atherosclerosis
Abstract
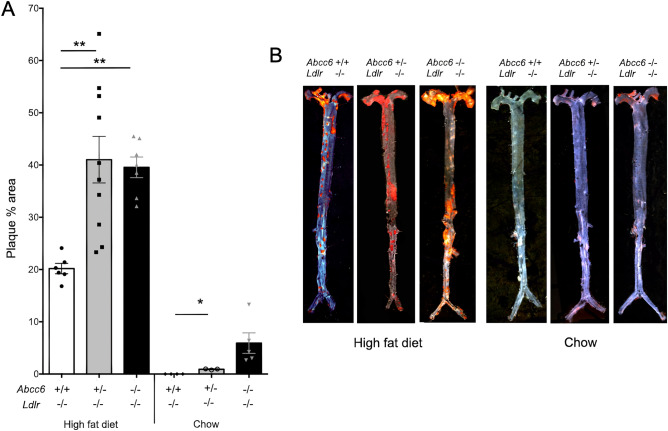
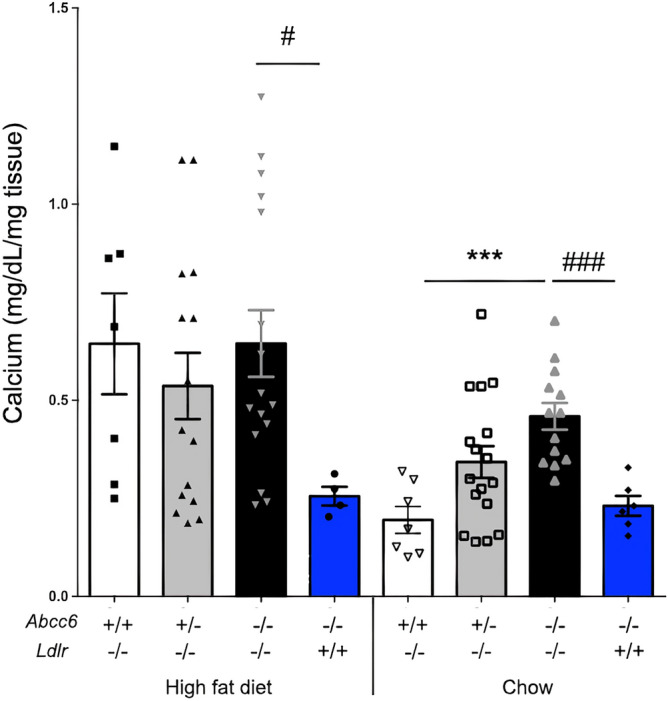
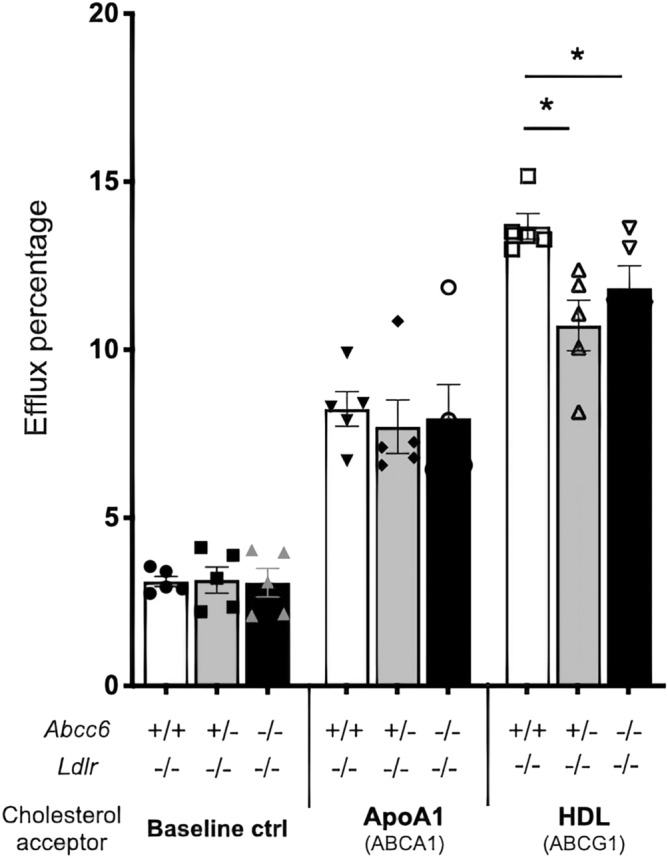
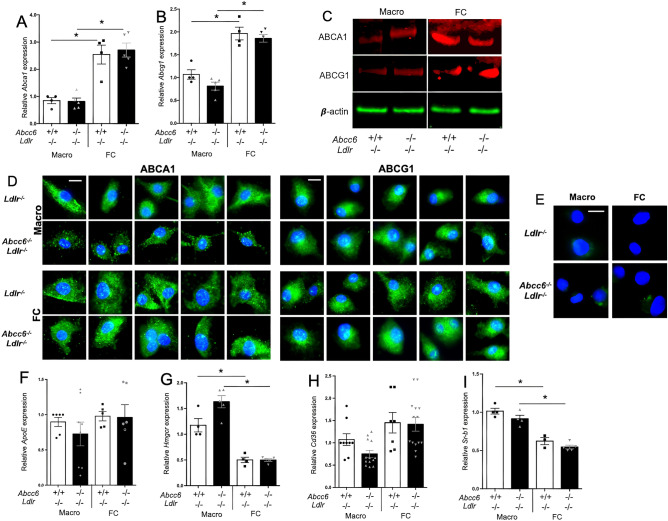
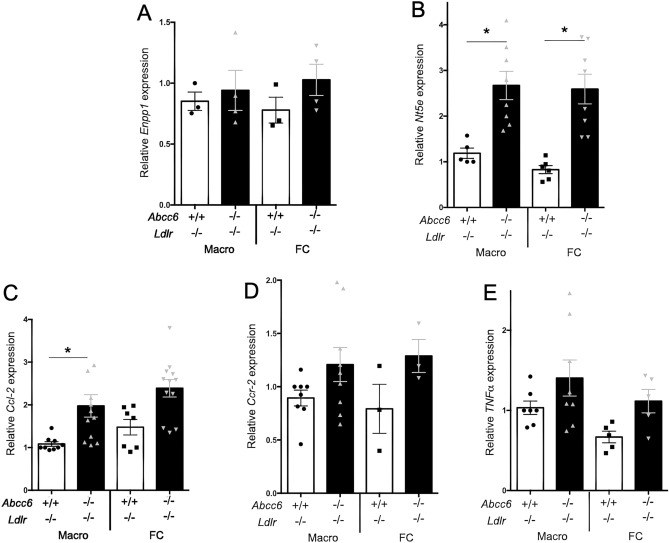
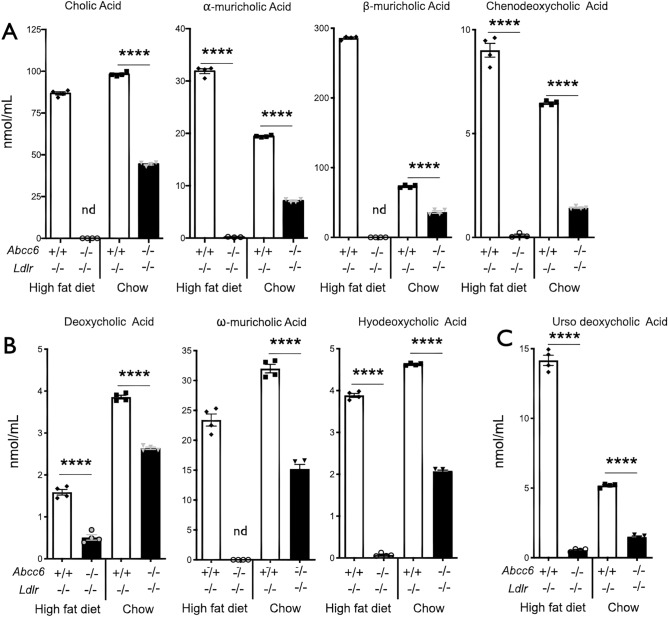
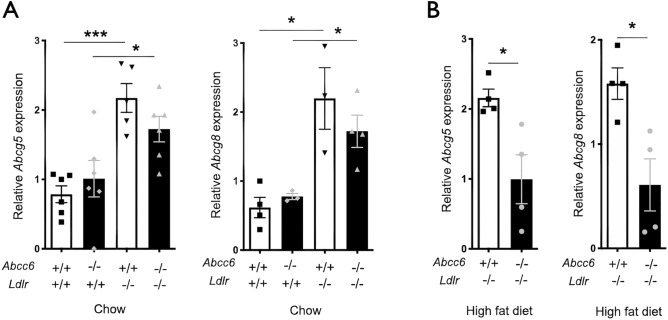
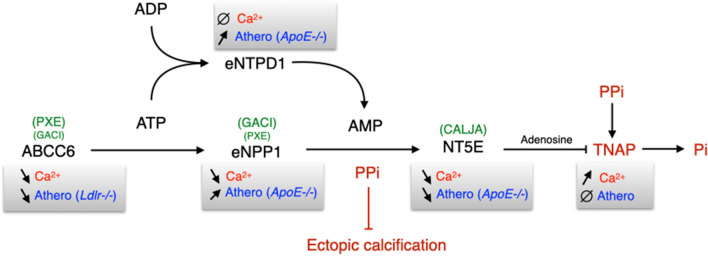
ABCC6 deficiency promotes ectopic calcification; however, circumstantial evidence suggested that ABCC6 may also influence atherosclerosis. The present study addressed the role of ABCC6 in atherosclerosis using Ldlr-/- mice and pseudoxanthoma elasticum (PXE) patients. Mice lacking the Abcc6 and Ldlr genes were fed an atherogenic diet for 16 weeks before intimal calcification, aortic plaque formation and lipoprotein profile were evaluated. Cholesterol efflux and the expression of several inflammation, atherosclerosis and cholesterol homeostasis-related genes were also determined in murine liver and bone marrow-derived macrophages. Furthermore, we examined plasma lipoproteins, vascular calcification, carotid intima-media thickness and atherosclerosis in a cohort of PXE patients with ABCC6 mutations and compared results to dysmetabolic subjects with increased cardiovascular risk. We found that ABCC6 deficiency causes changes in lipoproteins, with decreased HDL cholesterol in both mice and humans, and induces atherosclerosis. However, we found that the absence of ABCC6 does not influence overall vascular mineralization induced with atherosclerosis. Decreased cholesterol efflux from macrophage cells and other molecular changes such as increased pro-inflammation seen in both humans and mice are likely contributors for the phenotype. However, it is likely that other cellular and/or molecular mechanisms are involved. Our study showed a novel physiological role for ABCC6, influencing plasma lipoproteins and atherosclerosis in a haploinsufficient manner, with significant penetrance.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
