

Roles for ELMOD2 and Rootletin in ciliogenesis
- PMID: 33596093
- PMCID: PMC8108518
- DOI: 10.1091/mbc.E20-10-0635
Roles for ELMOD2 and Rootletin in ciliogenesis
Abstract
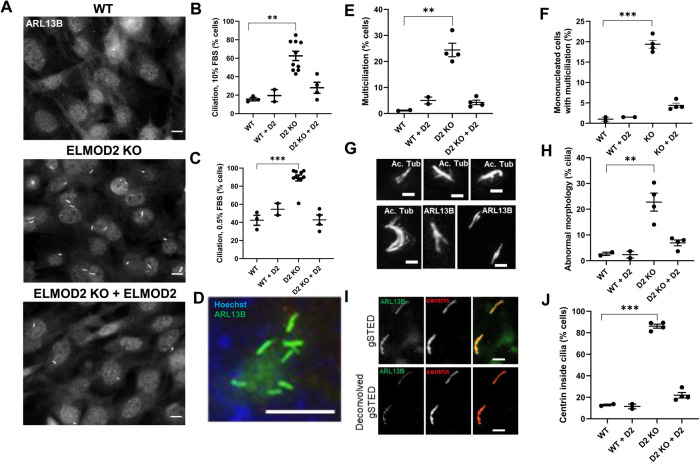
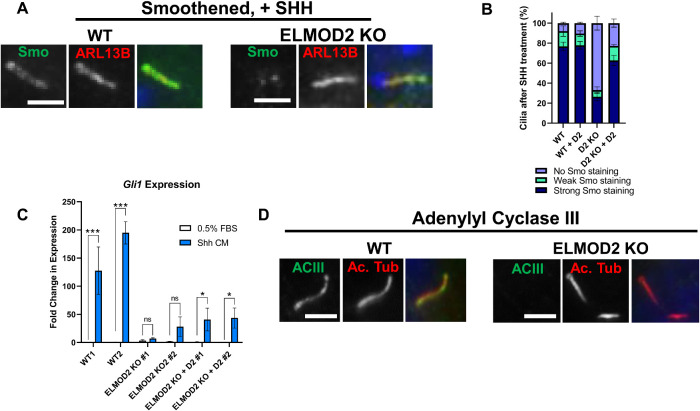
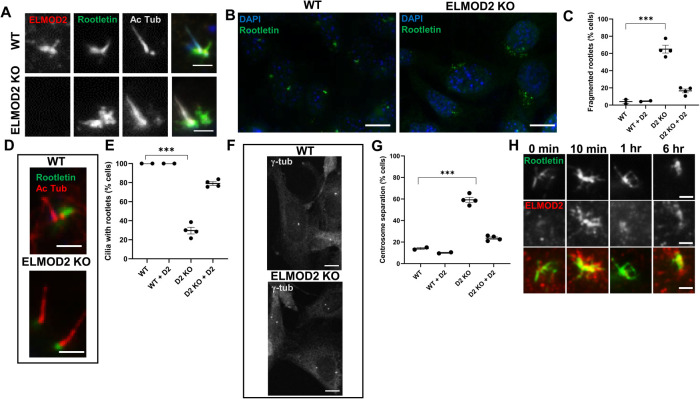
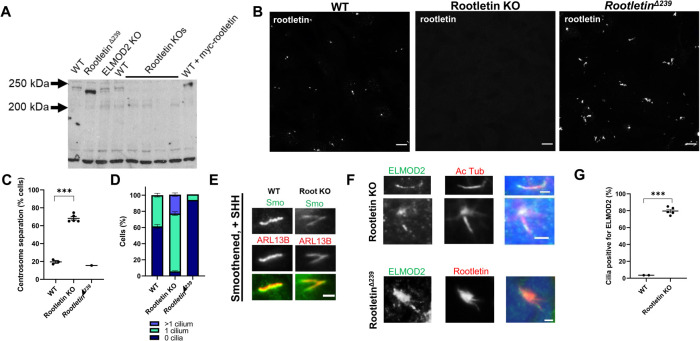
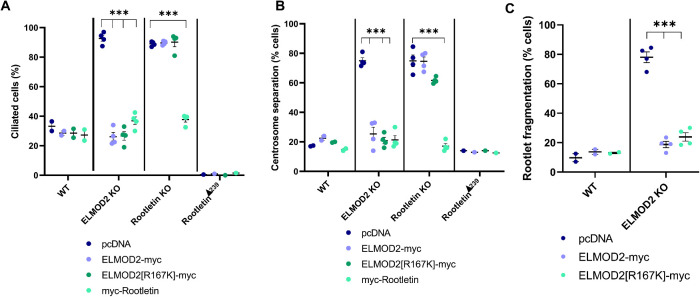
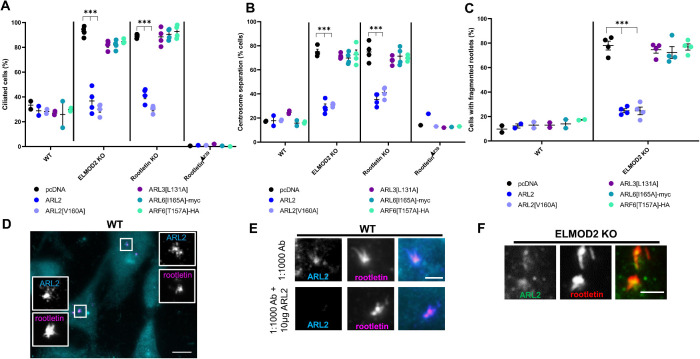
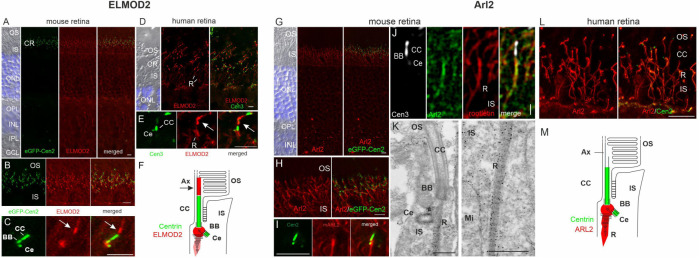
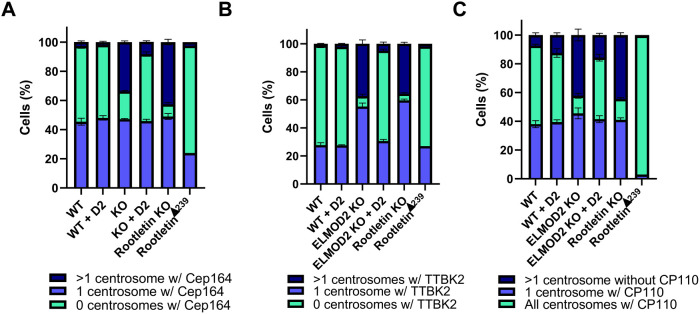
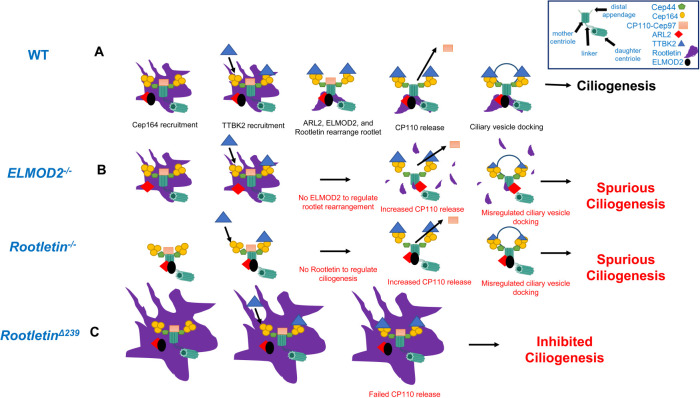
ELMOD2 is a GTPase-activating protein with uniquely broad specificity for ARF family GTPases. We previously showed that it acts with ARL2 in mitochondrial fusion and microtubule stability and with ARF6 during cytokinesis. Mouse embryonic fibroblasts deleted for ELMOD2 also displayed changes in cilia-related processes including increased ciliation, multiciliation, ciliary morphology, ciliary signaling, centrin accumulation inside cilia, and loss of rootlets at centrosomes with loss of centrosome cohesion. Increasing ARL2 activity or overexpressing Rootletin reversed these defects, revealing close functional links between the three proteins. This was further supported by the findings that deletion of Rootletin yielded similar phenotypes, which were rescued upon increasing ARL2 activity but not ELMOD2 overexpression. Thus, we propose that ARL2, ELMOD2, and Rootletin all act in a common pathway that suppresses spurious ciliation and maintains centrosome cohesion. Screening a number of markers of steps in the ciliation pathway supports a model in which ELMOD2, Rootletin, and ARL2 act downstream of TTBK2 and upstream of CP110 to prevent spurious release of CP110 and to regulate ciliary vesicle docking. These data thus provide evidence supporting roles for ELMOD2, Rootletin, and ARL2 in the regulation of ciliary licensing.
Figures
Similar articles
-
The ARF GAPs ELMOD1 and ELMOD3 act at the Golgi and cilia to regulate ciliogenesis and ciliary protein traffic.Mol Biol Cell. 2022 Feb 1;33(2):ar13. doi: 10.1091/mbc.E21-09-0443. Epub 2021 Nov 24. Mol Biol Cell. 2022. PMID: 34818063 Free PMC article.
-
The ARF GAP ELMOD2 acts with different GTPases to regulate centrosomal microtubule nucleation and cytokinesis.Mol Biol Cell. 2020 Aug 15;31(18):2070-2091. doi: 10.1091/mbc.E20-01-0012. Epub 2020 Jul 2. Mol Biol Cell. 2020. PMID: 32614697 Free PMC article.
-
ELMOD2 regulates mitochondrial fusion in a mitofusin-dependent manner, downstream of ARL2.Mol Biol Cell. 2019 May 1;30(10):1198-1213. doi: 10.1091/mbc.E18-12-0804. Epub 2019 Mar 13. Mol Biol Cell. 2019. PMID: 30865555 Free PMC article.
-
Higher order signaling: ARL2 as regulator of both mitochondrial fusion and microtubule dynamics allows integration of 2 essential cell functions.Small GTPases. 2016 Oct;7(4):188-196. doi: 10.1080/21541248.2016.1211069. Epub 2016 Jul 11. Small GTPases. 2016. PMID: 27400436 Free PMC article. Review.
-
ARF family GTPases with links to cilia.Am J Physiol Cell Physiol. 2020 Aug 1;319(2):C404-C418. doi: 10.1152/ajpcell.00188.2020. Epub 2020 Jun 10. Am J Physiol Cell Physiol. 2020. PMID: 32520609 Free PMC article. Review.
Cited by
-
ELMOD3-Rab1A-Flotillin2 cascade regulates lumen formation via vesicle trafficking in Ciona notochord.Open Biol. 2023 Mar;13(3):220367. doi: 10.1098/rsob.220367. Epub 2023 Mar 15. Open Biol. 2023. PMID: 36918025 Free PMC article.
-
Phylogenetic profiling and cellular analyses of ARL16 reveal roles in traffic of IFT140 and INPP5E.Mol Biol Cell. 2022 Apr 1;33(4):ar33. doi: 10.1091/mbc.E21-10-0509-T. Epub 2022 Feb 23. Mol Biol Cell. 2022. PMID: 35196065 Free PMC article.
-
Arf-like Protein 2 (ARL2) Controls Microtubule Neogenesis during Early Postnatal Photoreceptor Development.Cells. 2022 Dec 30;12(1):147. doi: 10.3390/cells12010147. Cells. 2022. PMID: 36611941 Free PMC article.
-
The ARF GAPs ELMOD1 and ELMOD3 act at the Golgi and cilia to regulate ciliogenesis and ciliary protein traffic.Mol Biol Cell. 2022 Feb 1;33(2):ar13. doi: 10.1091/mbc.E21-09-0443. Epub 2021 Nov 24. Mol Biol Cell. 2022. PMID: 34818063 Free PMC article.
-
The structure and function of centriolar rootlets.J Cell Sci. 2021 Aug 15;134(16):jcs258544. doi: 10.1242/jcs.258544. Epub 2021 Aug 18. J Cell Sci. 2021. PMID: 34405868 Free PMC article.
References
-
- Ahmadian MR, Stege P, Scheffzek K, Wittinghofer A (1997). Confirmation of the arginine-finger hypothesis for the GAP-stimulated GTP-hydrolysis reaction of Ras. Nat Struct Biol 4, 686–689. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
