

Intrinsic and chemically-induced daughter number variations in cancer cell lines
- PMID: 33596747
- PMCID: PMC8018351
- DOI: 10.1080/15384101.2021.1883363
Intrinsic and chemically-induced daughter number variations in cancer cell lines
Abstract
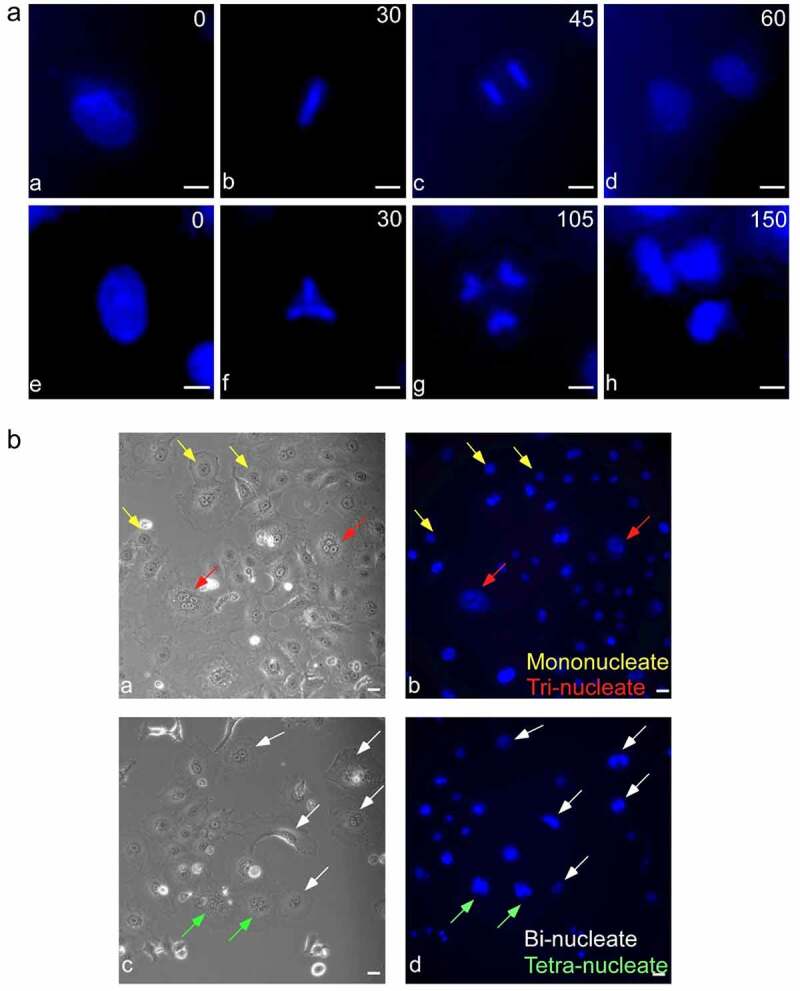
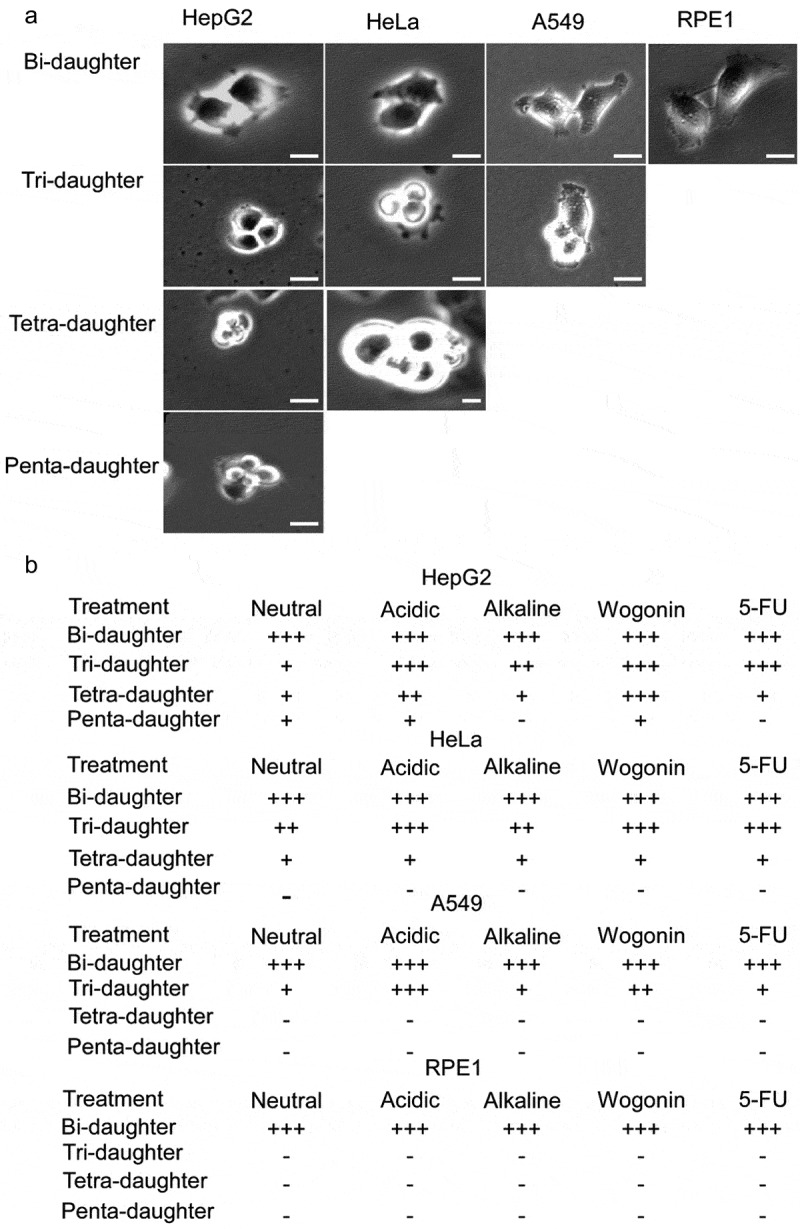
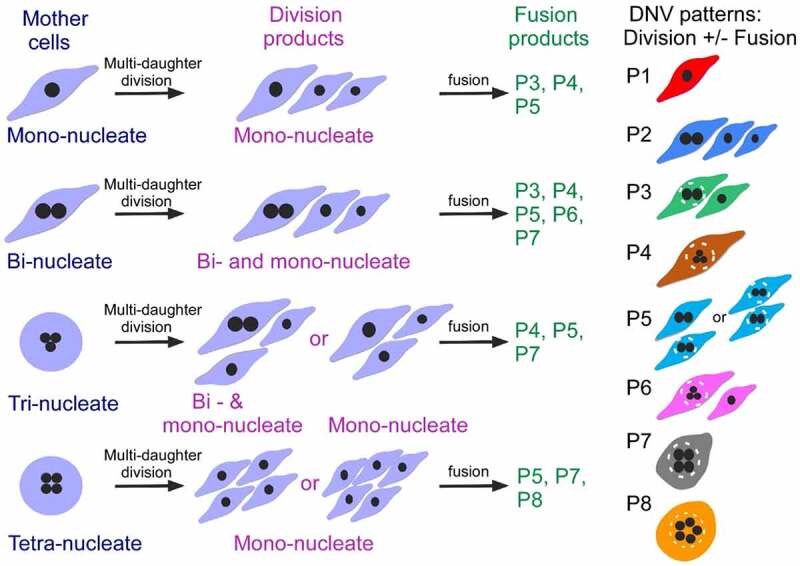
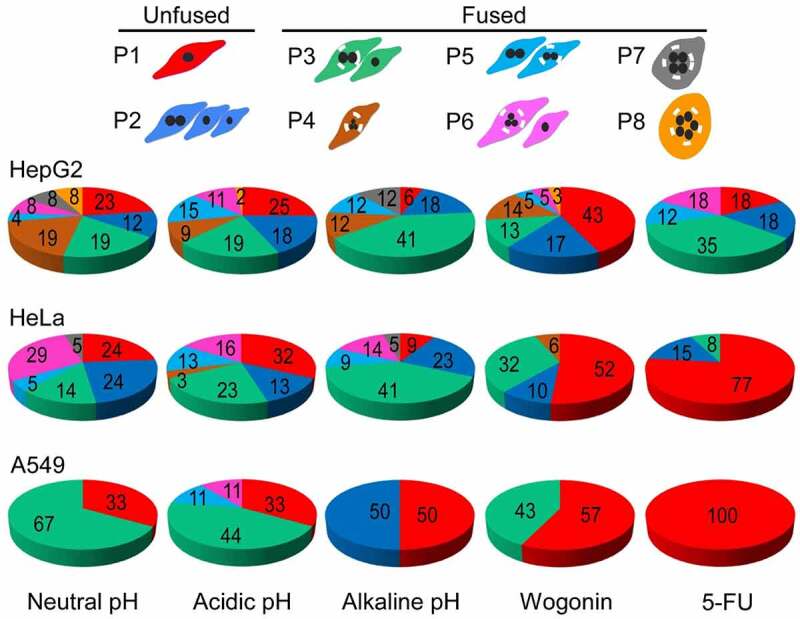
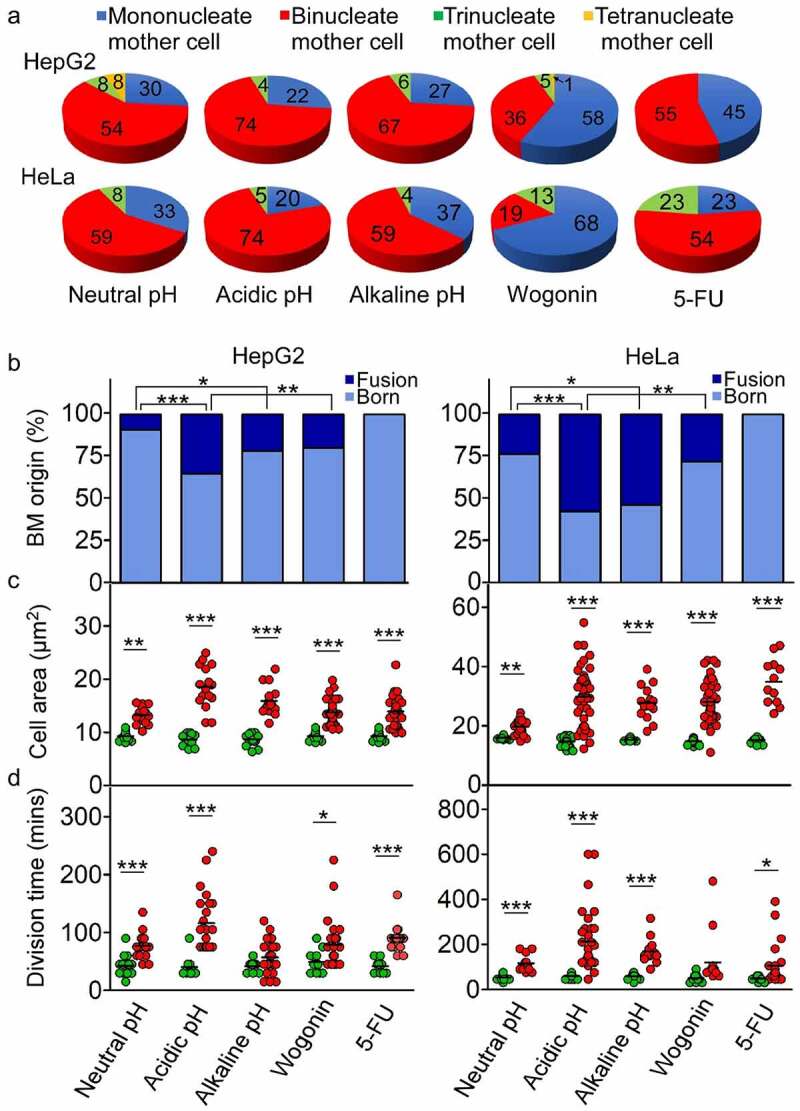
Multipolar mitosis was observed in cancer cells under mechanical stress or drug treatment. However, a comprehensive understanding of its basic properties and significance to cancer cell biology is lacking. In the present study, live-cell imaging was employed to investigate the division and nucleation patterns in four different cell lines. Multi-daughter divisions were observed in the three cancer cell lines HepG2, HeLa and A549, but not in the transformed non-cancer cell line RPE1. Multi-daughter mother cells displayed multi-nucleation, enlarged cell area, and prolonged division time. Under acidic pH or treatment with the anti-cancer drug 5-fluorouracil (5-FU) or the phytochemical compound wogonin, multi-daughter mitoses were increased to different extents in all three cancer cell lines, reaching as high as 16% of all mitoses. While less than 0.4% of the bi-daughter mitosis were followed by cell fusion events under the various treatment conditions, 50% or more of the multi-daughter mitoses were followed by fusion events at neutral, acidic or alkaline pH. These findings revealed a "Daughter Number Variation" (DNV) process in the cancer cells, with multi-daughter divisions in Stage 1 and cell fusions leading to the formation of cells containing up to five nuclei in Stage 2. The Stage 2-fusions were inhibited by 5-FU in A549 and HeLa, and by wogonin in A549, HeLa and HepG2. The parallel relationship between DNV frequency and malignancy among the different cell lines suggests that the inclusion of anti-fusion agents exemplified by wogonin and 5-FU could be beneficial in combinatory cancer chemotherapies.
Keywords: 5-fluorouracil; Cancer; cell fusion; multi-daughter division; wogonin.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Gisselsson D. Classification of chromosome segregation errors in cancer. Chromosoma. 2008;117(6):511–519. - PubMed
-
- McClelland SE. Role of chromosomal instability in cancer progression. Endocr Relat Cancer. 2017;24(9):23–31. - PubMed
-
- Hansemann V. On asymmetric cell division in epithelial cancers and their biological significance. Arch Pathol Anat Physiol Klin Med. 1890;119(119):299–236. DOI: 10.1007/BF01882039. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical