

Genomic Background Governs Opposing Responses to Nalidixic Acid upon Megaplasmid Acquisition in Pseudomonas
- PMID: 33597171
- PMCID: PMC8544880
- DOI: 10.1128/mSphere.00008-21
Genomic Background Governs Opposing Responses to Nalidixic Acid upon Megaplasmid Acquisition in Pseudomonas
Abstract
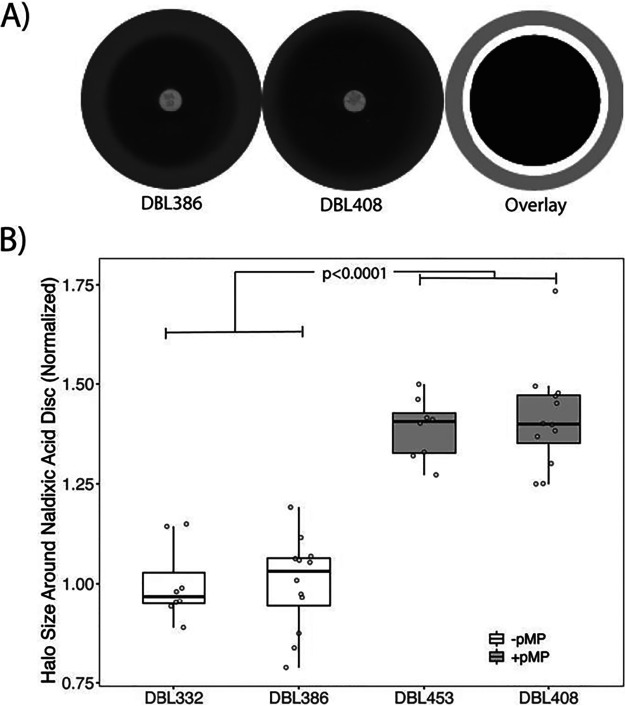
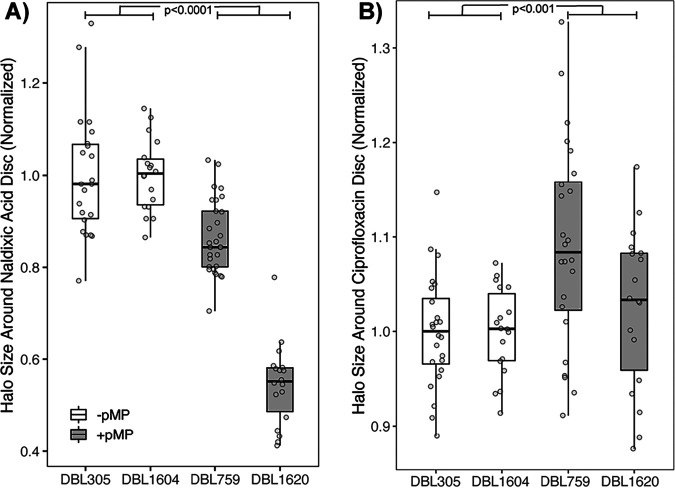
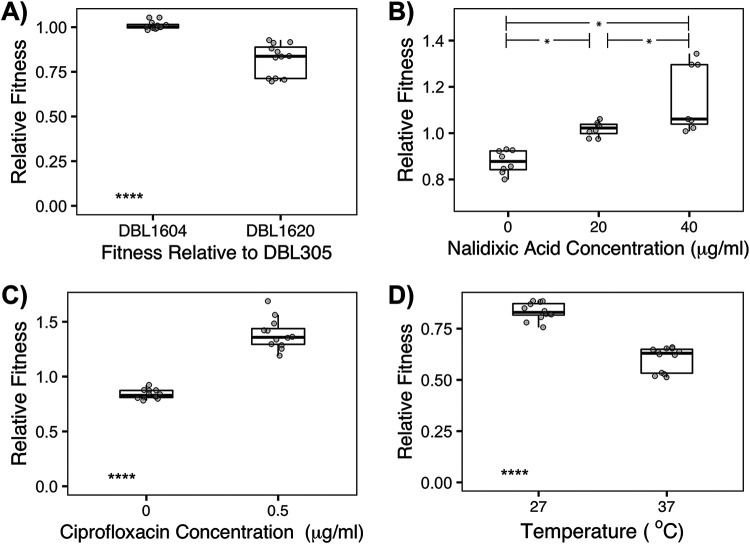
Horizontal gene transfer is a significant driver of evolutionary dynamics across microbial populations. Although the benefits of the acquisition of new genetic material are often quite clear, experiments across systems have demonstrated that gene transfer events can cause significant phenotypic changes and entail fitness costs in a way that is dependent on the genomic and environmental context. Here, we test for the generality of one previously identified cost, sensitization of cells to the antibiotic nalidixic acid after acquisition of an ∼1-Mb megaplasmid, across Pseudomonas strains and species. Overall, we find that the presence of this megaplasmid sensitizes many different Pseudomonas strains to nalidixic acid but that this same horizontal gene transfer event increases resistance of Pseudomonas putida KT2440 to nalidixic acid across assays as well as to ciprofloxacin under competitive conditions. These phenotypic results are not easily explained away as secondary consequences of overall fitness effects and appear to occur independently of another cost associated with this megaplasmid, sensitization to higher temperatures. Lastly, we draw parallels between these reported results and the phenomenon of sign epistasis for de novo mutations and explore how context dependence of effects of plasmid acquisition could impact overall evolutionary dynamics and the evolution of antimicrobial resistance.IMPORTANCE Numerous studies have demonstrated that gene transfer events (e.g., plasmid acquisition) can entail a variety of costs that arise as by-products of the incorporation of foreign DNA into established physiological and genetic systems. These costs can be ameliorated through evolutionary time by the occurrence of compensatory mutations, which stabilize the presence of a horizontally transferred region within the genome but which also may skew future adaptive possibilities for these lineages. Here, we demonstrate another possible outcome, that phenotypic changes arising as a consequence of the same horizontal gene transfer (HGT) event are costly to some strains but may actually be beneficial in other genomic backgrounds under the right conditions. These results provide a new viewpoint for considering conditions that promote plasmid maintenance and highlight the influence of genomic and environmental contexts when considering amelioration of fitness costs after HGT events.
Keywords: antibiotic resistance; drug resistance evolution; horizontal gene transfer; megaplasmids; quinolones.
Copyright © 2021 Baltrus et al.
Figures

References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
