

Neurodevelopmental defects and neurodegenerative phenotypes in human brain organoids carrying Parkinson's disease-linked DNAJC6 mutations
- PMID: 33597231
- PMCID: PMC7888924
- DOI: 10.1126/sciadv.abb1540
Neurodevelopmental defects and neurodegenerative phenotypes in human brain organoids carrying Parkinson's disease-linked DNAJC6 mutations
Erratum in
-
Erratum for the Research Article: "Neurodevelopmental defects and neurodegenerative phenotypes in human brain organoids carrying Parkinson's disease-linked DNAJC6 mutations".Sci Adv. 2023 Nov 3;9(44):eadl1498. doi: 10.1126/sciadv.adl1498. Epub 2023 Nov 3. Sci Adv. 2023. PMID: 37922364 Free PMC article. No abstract available.
Abstract
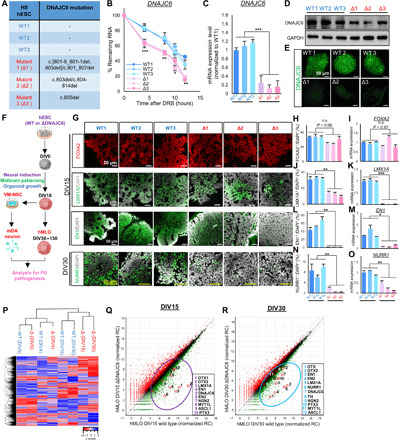
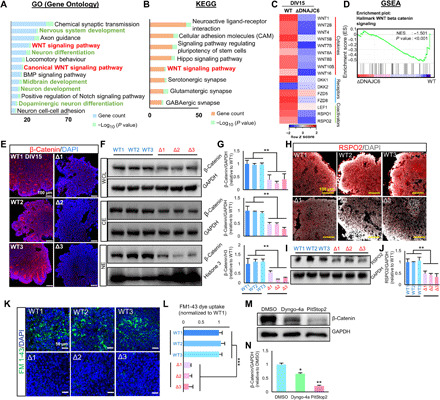
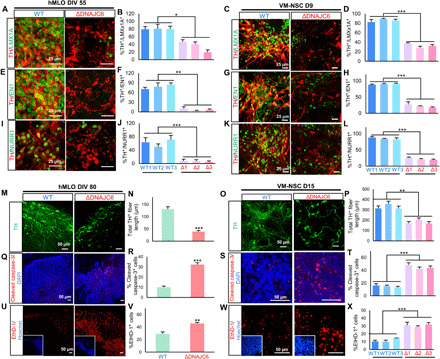
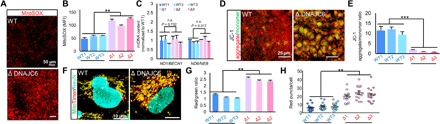
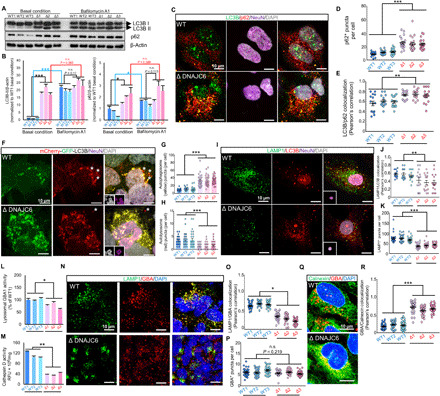
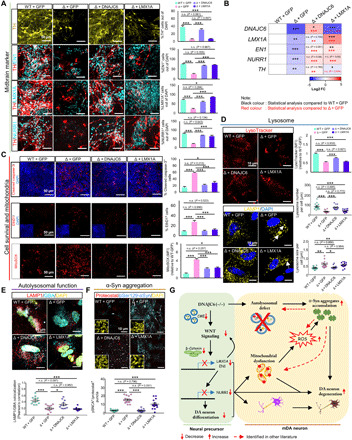
Loss-of-function mutations of DNAJC6, encoding HSP40 auxilin, have recently been identified in patients with early-onset Parkinson's disease (PD). To study the roles of DNAJC6 in PD pathogenesis, we used human embryonic stem cells with CRISPR-Cas9-mediated gene editing. Here, we show that DNAJC6 mutations cause key PD pathologic features, i.e., midbrain-type dopamine (mDA) neuron degeneration, pathologic α-synuclein aggregation, increase of intrinsic neuronal firing frequency, and mitochondrial and lysosomal dysfunctions in human midbrain-like organoids (hMLOs). In addition, neurodevelopmental defects were also manifested in hMLOs carrying the mutations. Transcriptomic analyses followed by experimental validation revealed that defects in DNAJC6-mediated endocytosis impair the WNT-LMX1A signal during the mDA neuron development. Furthermore, reduced LMX1A expression during development caused the generation of vulnerable mDA neurons with the pathologic manifestations. These results suggest that the human model of DNAJC6-PD recapitulates disease phenotypes and reveals mechanisms underlying disease pathology, providing a platform for assessing therapeutic interventions.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures

References
-
- Poewe W., Seppi K., Tanner C. M., Halliday G. M., Brundin P., Volkmann J., Schrag A. E., Lang A. E., Parkinson disease. Nat. Rev. Dis. Primers. 3, 17013 (2017). - PubMed
-
- Ungewickell E., Ungewickell H., Holstein S. E., Lindner R., Prasad K., Barouch W., Martin B., Greene L. E., Eisenberg E., Role of auxilin in uncoating clathrin-coated vesicles. Nature 378, 632–635 (1995). - PubMed
-
- Edvardson S., Cinnamon Y., Ta-Shma A., Shaag A., Yim Y. I., Zenvirt S., Jalas C., Lesage S., Brice A., Taraboulos A., Kaestner K. H., Greene L. E., Elpeleg O., A deleterious mutation in DNAJC6 encoding the neuronal-specific clathrin-uncoating co-chaperone auxilin, is associated with juvenile parkinsonism. PLOS ONE 7, e36458 (2012). - PMC - PubMed
-
- Koroglu C., Baysal L., Cetinkaya M., Karasoy H., Tolun A., DNAJC6 is responsible for juvenile parkinsonism with phenotypic variability. Parkinsonism Relat. Disord. 19, 320–324 (2013). - PubMed
-
- Olgiati S., Quadri M., Fang M., Rood J. P., Saute J. A., Chien H. F., Bouwkamp C. G., Graafland J., Minneboo M., Breedveld G. J., Zhang J.; International Parkinsonism Genetics Network, Verheijen F. W., Boon A. J., Kievit A. J., Jardim L. B., Mandemakers W., Barbosa E. R., Rieder C. R., Leenders K. L., Wang J., Bonifati V., DNAJC6 mutations associated with early-onset Parkinson’s disease. Ann. Neurol. 79, 244–256 (2016). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
