

An engineered decoy receptor for SARS-CoV-2 broadly binds protein S sequence variants
- PMID: 33597251
- PMCID: PMC7888922
- DOI: 10.1126/sciadv.abf1738
An engineered decoy receptor for SARS-CoV-2 broadly binds protein S sequence variants
Abstract
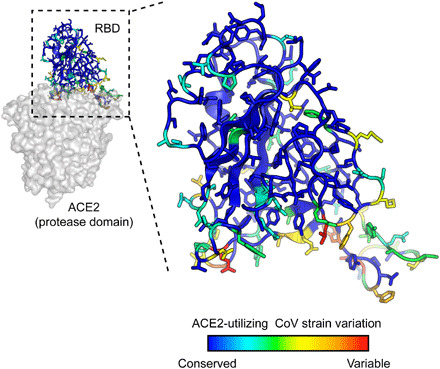
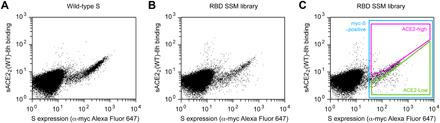
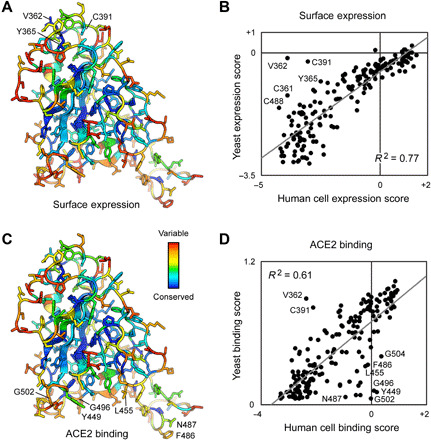
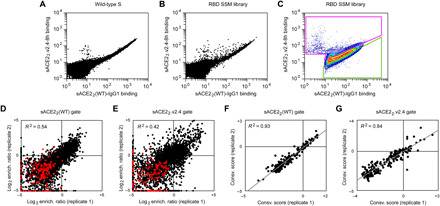
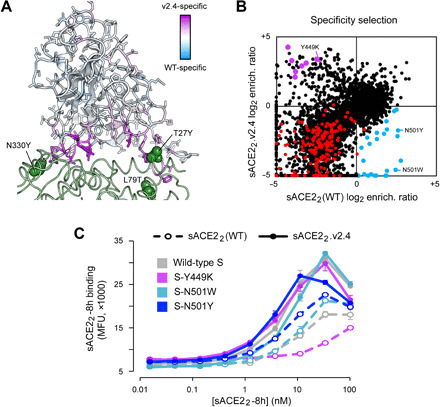
The spike S of SARS-CoV-2 recognizes ACE2 on the host cell membrane to initiate entry. Soluble decoy receptors, in which the ACE2 ectodomain is engineered to block S with high affinity, potently neutralize infection and, because of close similarity with the natural receptor, hold out the promise of being broadly active against virus variants without opportunity for escape. Here, we directly test this hypothesis. We find that an engineered decoy receptor, sACE22v2.4, tightly binds S of SARS-associated viruses from humans and bats, despite the ACE2-binding surface being a region of high diversity. Saturation mutagenesis of the receptor-binding domain followed by in vitro selection, with wild-type ACE2 and the engineered decoy competing for binding sites, failed to find S mutants that discriminate in favor of the wild-type receptor. We conclude that resistance to engineered decoys will be rare and that decoys may be active against future outbreaks of SARS-associated betacoronaviruses.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution License 4.0 (CC BY).
Figures
Update of
-
An engineered decoy receptor for SARS-CoV-2 broadly binds protein S sequence variants.bioRxiv [Preprint]. 2020 Dec 21:2020.10.18.344622. doi: 10.1101/2020.10.18.344622. bioRxiv. 2020. Update in: Sci Adv. 2021 Feb 17;7(8):eabf1738. doi: 10.1126/sciadv.abf1738. PMID: 33398275 Free PMC article. Updated. Preprint.
References
-
- Zhou P., Yang X.-L., Wang X.-G., Hu B., Zhang L., Zhang W., Si H.-R., Zhu Y., Li B., Huang C.-L., Chen H.-D., Chen J., Luo Y., Guo H., Jiang R.-D., Liu M.-Q., Chen Y., Shen X.-R., Wang X., Zheng X.-S., Zhao K., Chen Q.-J., Deng F., Liu L.-L., Yan B., Zhan F.-X., Wang Y.-Y., Xiao G.-F., Shi Z.-L., A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579, 270–273 (2020). - PMC - PubMed
-
- Hoffmann M., Kleine-Weber H., Schroeder S., Krüger N., Herrler T., Erichsen S., Schiergens T. S., Herrler G., Wu N.-H., Nitsche A., Müller M. A., Drosten C., Pöhlmann S., SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 181, 271–280.e8 (2020). - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
