

Molecular mechanisms of assembly and TRIP13-mediated remodeling of the human Shieldin complex
- PMID: 33597306
- PMCID: PMC7923543
- DOI: 10.1073/pnas.2024512118
Molecular mechanisms of assembly and TRIP13-mediated remodeling of the human Shieldin complex
Abstract
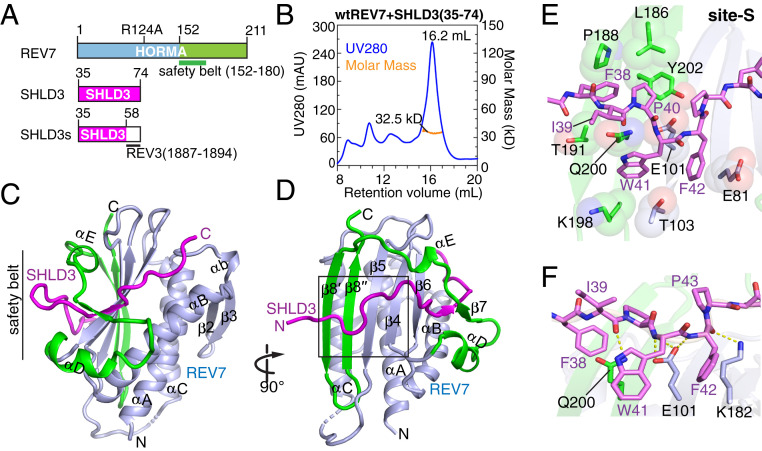
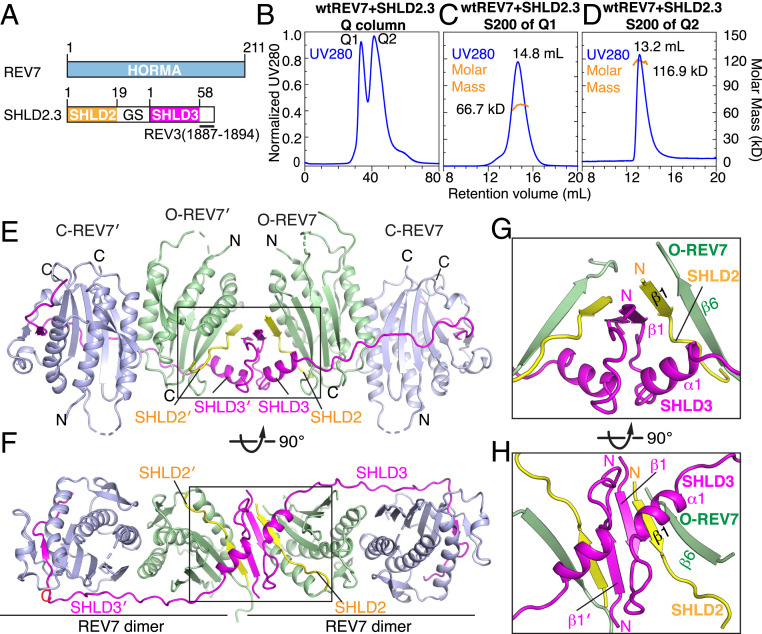
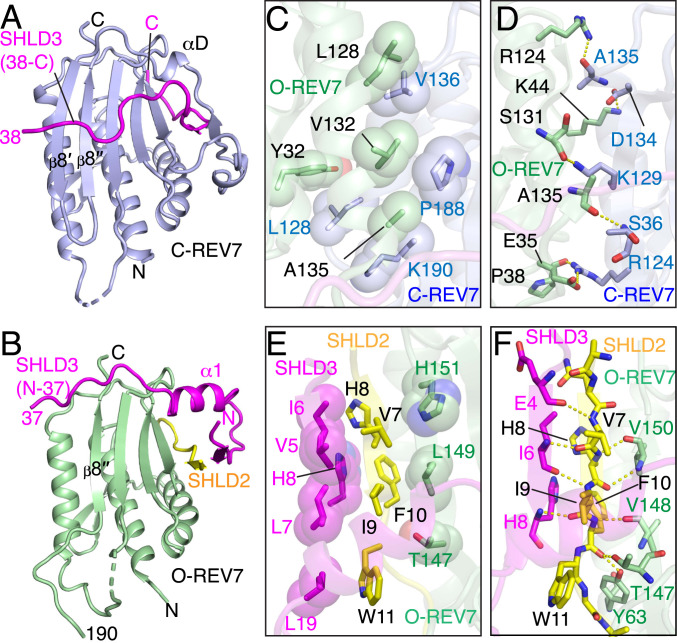
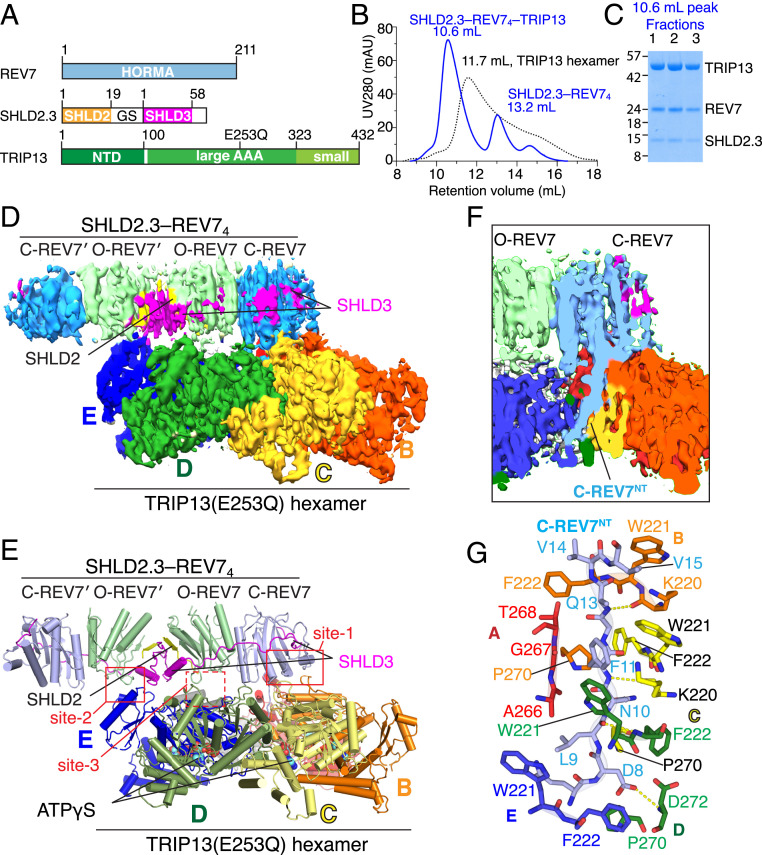
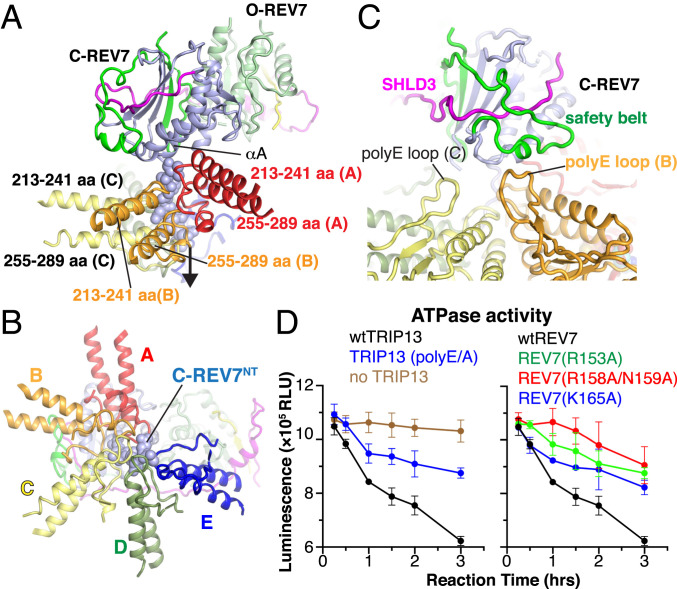
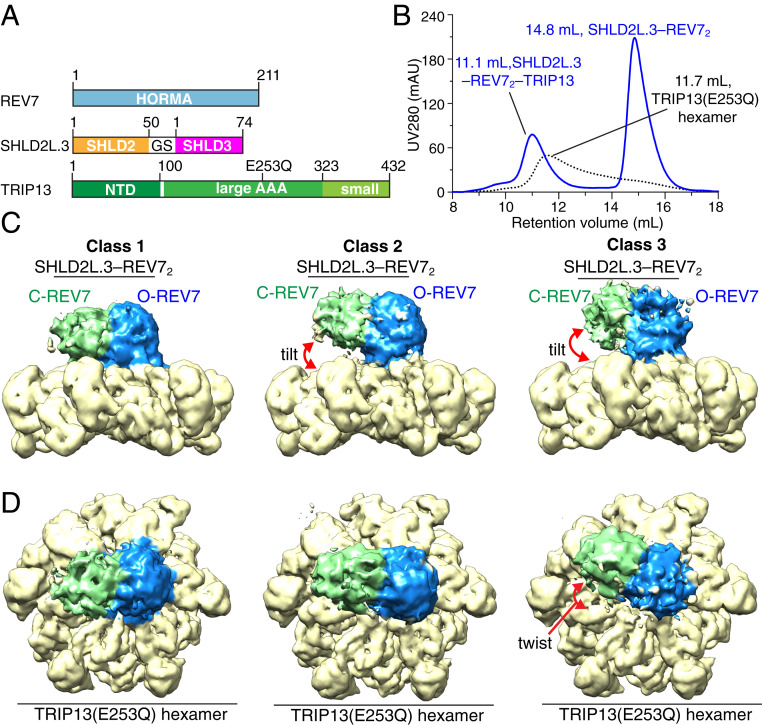
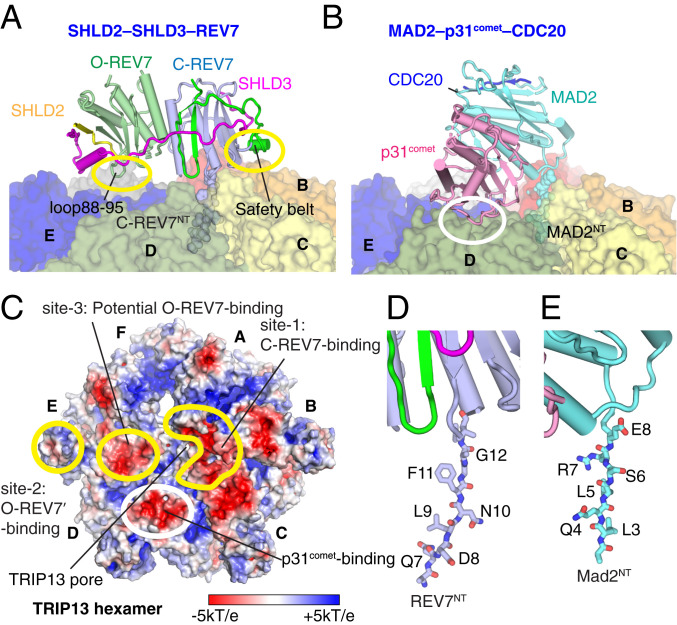
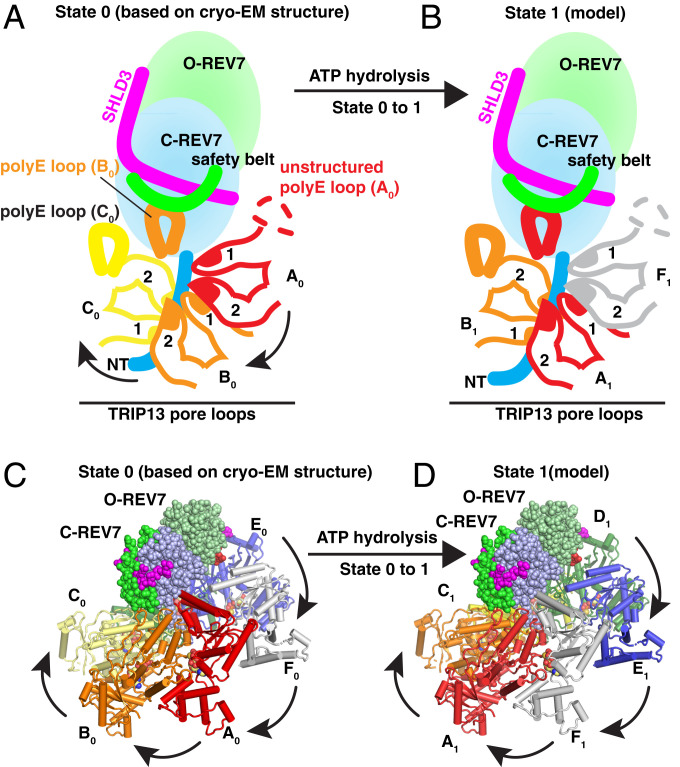
The Shieldin complex, composed of REV7, SHLD1, SHLD2, and SHLD3, protects DNA double-strand breaks (DSBs) to promote nonhomologous end joining. The AAA+ ATPase TRIP13 remodels Shieldin to regulate DNA repair pathway choice. Here we report crystal structures of human SHLD3-REV7 binary and fused SHLD2-SHLD3-REV7 ternary complexes, revealing that assembly of Shieldin requires fused SHLD2-SHLD3 induced conformational heterodimerization of open (O-REV7) and closed (C-REV7) forms of REV7. We also report the cryogenic electron microscopy (cryo-EM) structures of the ATPγS-bound fused SHLD2-SHLD3-REV7-TRIP13 complexes, uncovering the principles underlying the TRIP13-mediated disassembly mechanism of the Shieldin complex. We demonstrate that the N terminus of REV7 inserts into the central channel of TRIP13, setting the stage for pulling the unfolded N-terminal peptide of C-REV7 through the central TRIP13 hexameric channel. The primary interface involves contacts between the safety-belt segment of C-REV7 and a conserved and negatively charged loop of TRIP13. This process is mediated by ATP hydrolysis-triggered rotatory motions of the TRIP13 ATPase, thereby resulting in the disassembly of the Shieldin complex.
Keywords: SHLD2–SHLD3–REV7 complex; SHLD2–SHLD3–REV7–TRIP13 complex; SHLD3–REV7 complex; Shieldin assembly; TRIP13-mediated disassembly of Shieldin.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Khanna K. K., Jackson S. P., DNA double-strand breaks: Signaling, repair and the cancer connection. Nat. Genet. 27, 247–254 (2001). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
