

The structural basis of function and regulation of neuronal cotransporters NKCC1 and KCC2
- PMID: 33597714
- PMCID: PMC7889885
- DOI: 10.1038/s42003-021-01750-w
The structural basis of function and regulation of neuronal cotransporters NKCC1 and KCC2
Abstract
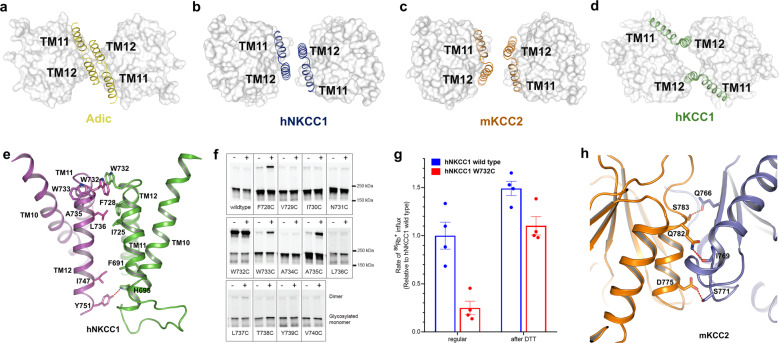
NKCC and KCC transporters mediate coupled transport of Na++K++Cl- and K++Cl- across the plasma membrane, thus regulating cell Cl- concentration and cell volume and playing critical roles in transepithelial salt and water transport and in neuronal excitability. The function of these transporters has been intensively studied, but a mechanistic understanding has awaited structural studies of the transporters. Here, we present the cryo-electron microscopy (cryo-EM) structures of the two neuronal cation-chloride cotransporters human NKCC1 (SLC12A2) and mouse KCC2 (SLC12A5), along with computational analysis and functional characterization. These structures highlight essential residues in ion transport and allow us to propose mechanisms by which phosphorylation regulates transport activity.
Conflict of interest statement
The authors declare no competing interests.
Figures

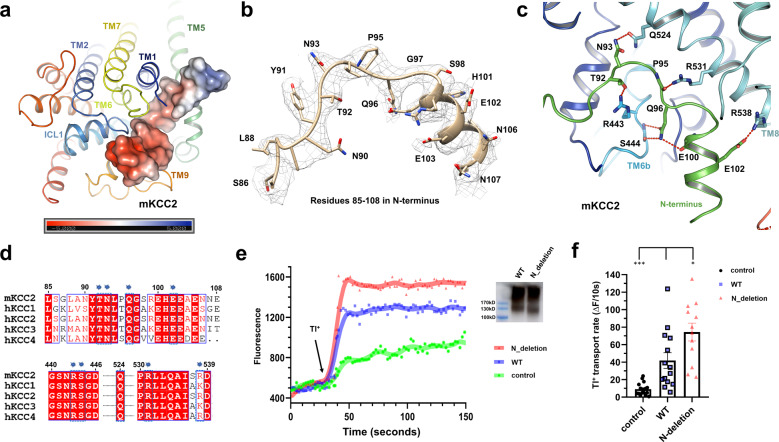
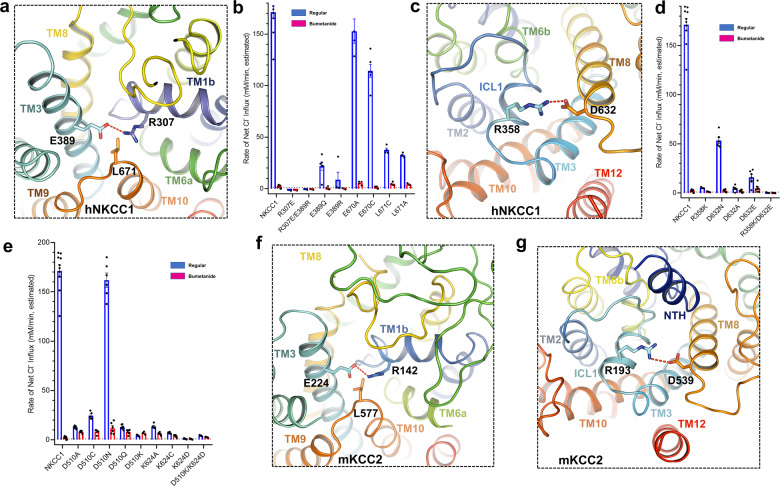
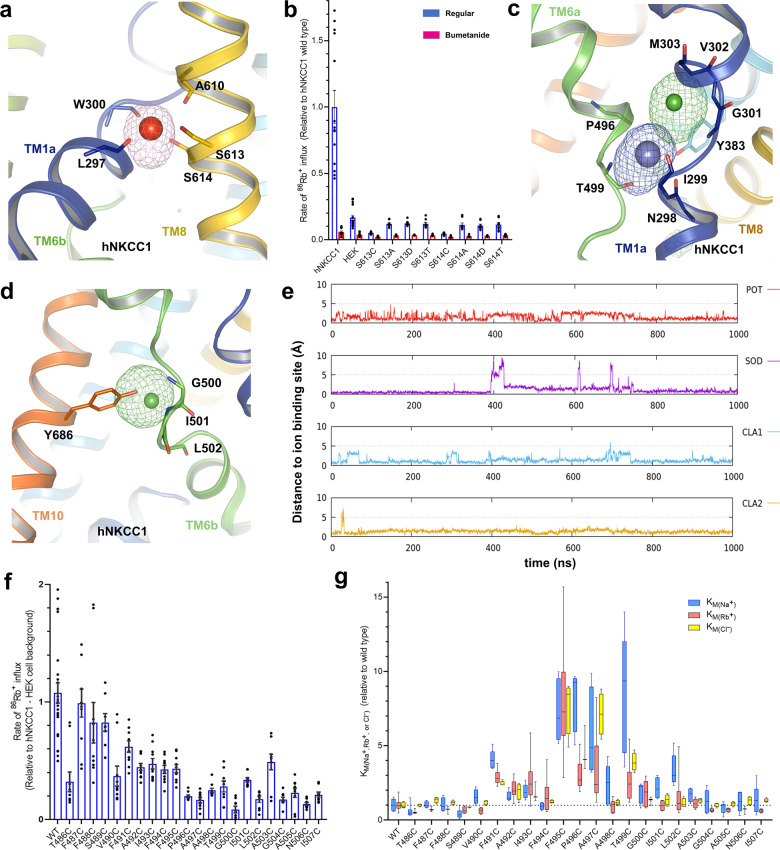
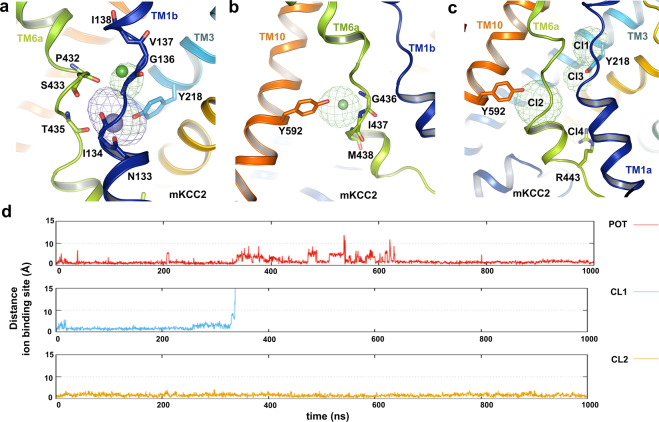

Similar articles
-
High-Resolution Views and Transport Mechanisms of the NKCC1 and KCC Transporters.J Mol Biol. 2021 Aug 6;433(16):167056. doi: 10.1016/j.jmb.2021.167056. Epub 2021 May 20. J Mol Biol. 2021. PMID: 34022207 Free PMC article. Review.
-
Structure and mechanism of the cation-chloride cotransporter NKCC1.Nature. 2019 Aug;572(7770):488-492. doi: 10.1038/s41586-019-1438-2. Epub 2019 Jul 31. Nature. 2019. PMID: 31367042 Free PMC article.
-
Structure of the human cation-chloride cotransporter NKCC1 determined by single-particle electron cryo-microscopy.Nat Commun. 2020 Feb 21;11(1):1016. doi: 10.1038/s41467-020-14790-3. Nat Commun. 2020. PMID: 32081947 Free PMC article.
-
All-Atom Simulations Uncover the Molecular Terms of the NKCC1 Transport Mechanism.J Chem Inf Model. 2021 Jul 26;61(7):3649-3658. doi: 10.1021/acs.jcim.1c00551. Epub 2021 Jul 2. J Chem Inf Model. 2021. PMID: 34213892
-
SLC12A cryo-EM: analysis of relevant ion binding sites, structural domains, and amino acids.Am J Physiol Cell Physiol. 2023 Oct 1;325(4):C921-C939. doi: 10.1152/ajpcell.00089.2023. Epub 2023 Aug 7. Am J Physiol Cell Physiol. 2023. PMID: 37545407 Review.
Cited by
-
The choroid plexus: a missing link in our understanding of brain development and function.Physiol Rev. 2023 Jan 1;103(1):919-956. doi: 10.1152/physrev.00060.2021. Epub 2022 Sep 29. Physiol Rev. 2023. PMID: 36173801 Free PMC article. Review.
-
Role of NKCC1 and KCC2 during hypoxia-induced neuronal swelling in the neonatal neocortex.Neurobiol Dis. 2023 Mar;178:106013. doi: 10.1016/j.nbd.2023.106013. Epub 2023 Jan 25. Neurobiol Dis. 2023. PMID: 36706928 Free PMC article.
-
A Futile Cycle?: Tissue Homeostatic Trans-Membrane Water Co-Transport: Kinetics, Thermodynamics, Metabolic Consequences.bioRxiv [Preprint]. 2024 Apr 26:2024.04.17.589812. doi: 10.1101/2024.04.17.589812. bioRxiv. 2024. PMID: 38659823 Free PMC article. Preprint.
-
The biogenesis of potassium transporters: implications of disease-associated mutations.Crit Rev Biochem Mol Biol. 2024 Jun-Aug;59(3-4):154-198. doi: 10.1080/10409238.2024.2369986. Epub 2024 Jul 1. Crit Rev Biochem Mol Biol. 2024. PMID: 38946646 Free PMC article. Review.
-
Converging Mechanisms of Epileptogenesis and Their Insight in Glioblastoma.Front Mol Neurosci. 2022 Jun 27;15:903115. doi: 10.3389/fnmol.2022.903115. eCollection 2022. Front Mol Neurosci. 2022. PMID: 35832394 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
