

ALMS1 Regulates TGF-β Signaling and Morphology of Primary Cilia
- PMID: 33598462
- PMCID: PMC7882606
- DOI: 10.3389/fcell.2021.623829
ALMS1 Regulates TGF-β Signaling and Morphology of Primary Cilia
Abstract
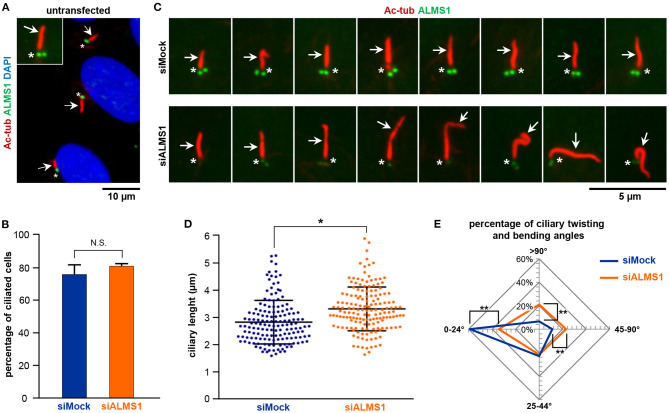
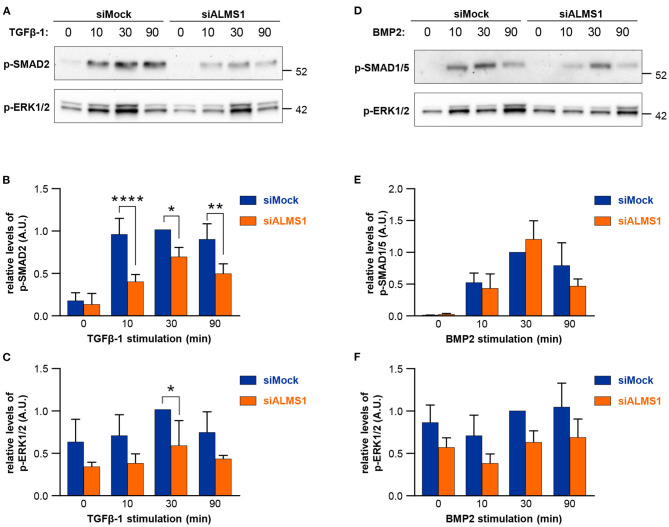
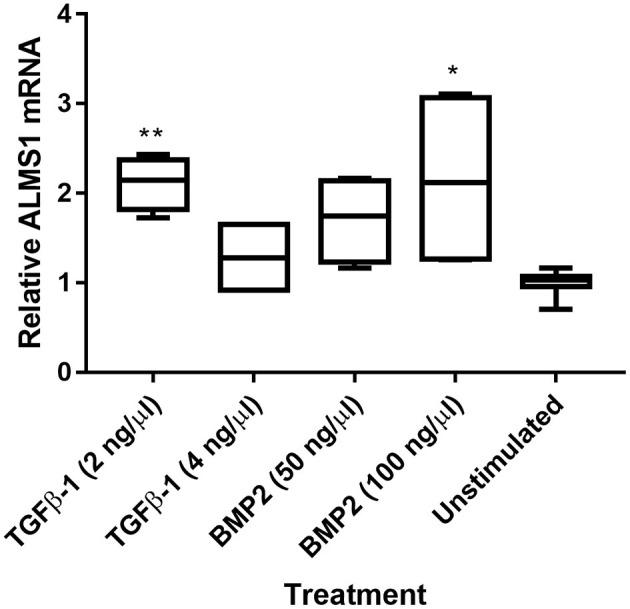
In this study, we aimed to evaluate the role of ALMS1 in the morphology of primary cilia and regulation of cellular signaling using a knockdown model of the hTERT-RPE1 cell line. ALMS1 depletion resulted in the formation of longer cilia, which often displayed altered morphology as evidenced by extensive twisting and bending of the axoneme. Transforming growth factor beta/bone morphogenetic protein (TGF-β/BMP) signaling, which is regulated by primary cilia, was similarly affected by ALMS1 depletion as judged by reduced levels of TGFβ-1-mediated activation of SMAD2/3. These results provide novel information on the role of ALMS1 in the function of primary cilia and processing of cellular signaling, which when aberrantly regulated may underlie Alström syndrome.
Keywords: ALMS1; Alström syndrome (AS); TGF-β/BMP signaling; ciliary length; ciliary morphology; ciliopathies; hTERT RPE-1 cells; primary cilium.
Copyright © 2021 Álvarez-Satta, Lago-Docampo, Bea-Mascato, Solarat, Castro-Sánchez, Christensen and Valverde.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources