

The Structural and Biochemical Basis of Apocarotenoid Processing by β-Carotene Oxygenase-2
- PMID: 33600157
- PMCID: PMC8082539
- DOI: 10.1021/acschembio.0c00832
The Structural and Biochemical Basis of Apocarotenoid Processing by β-Carotene Oxygenase-2
Abstract
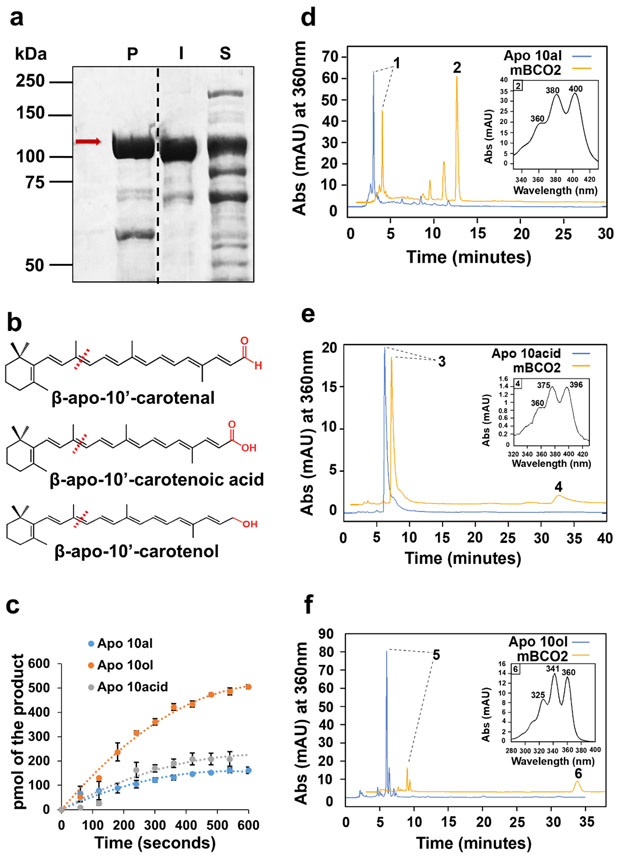
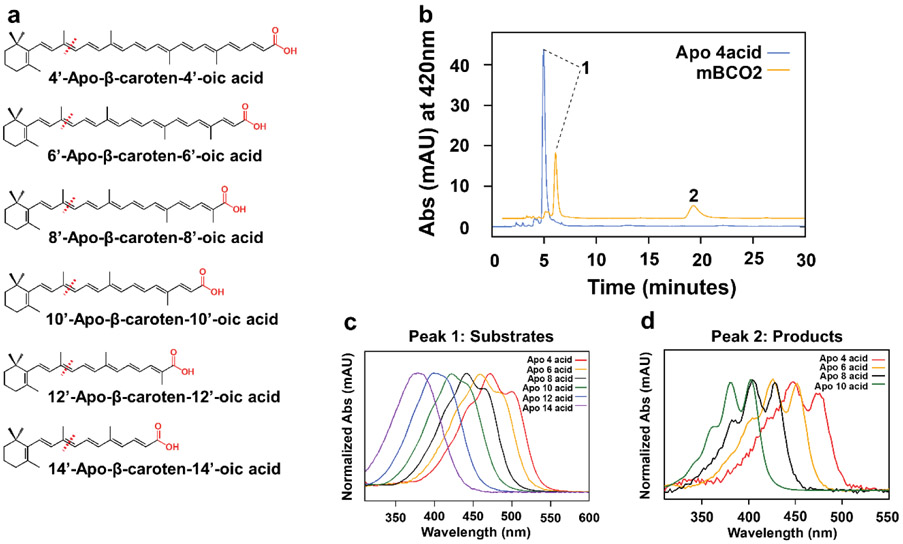
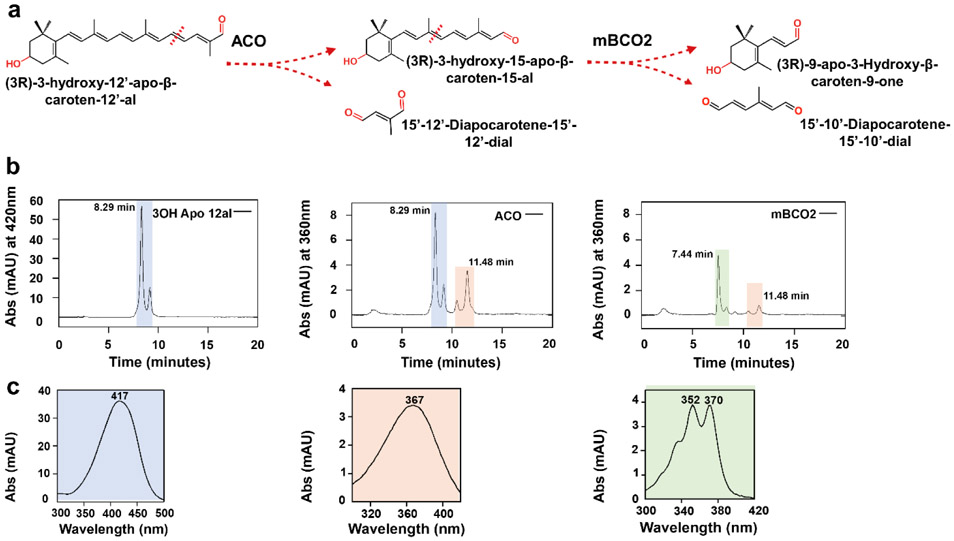
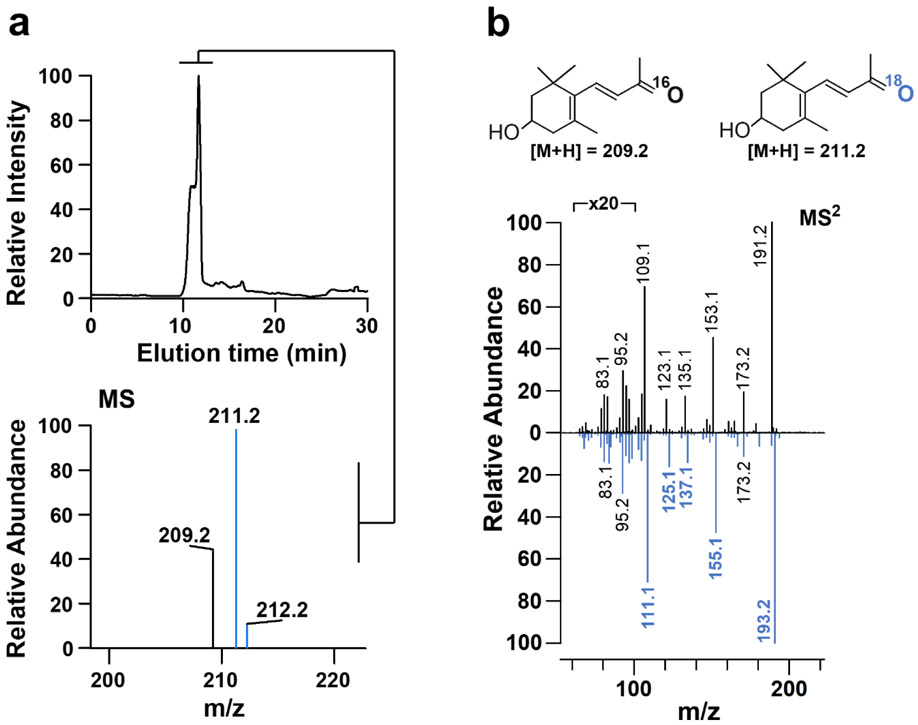
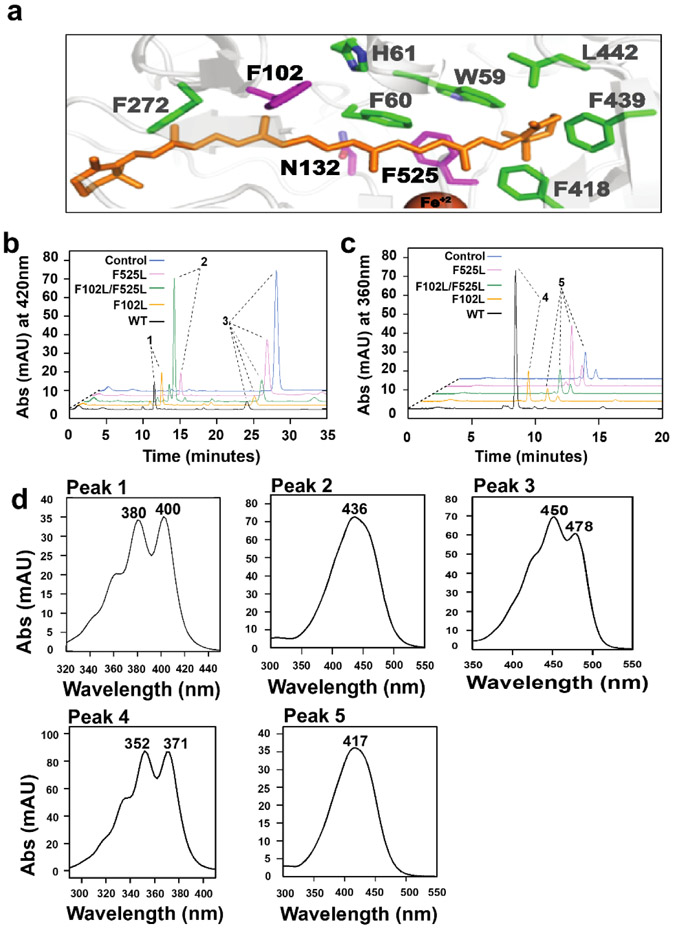
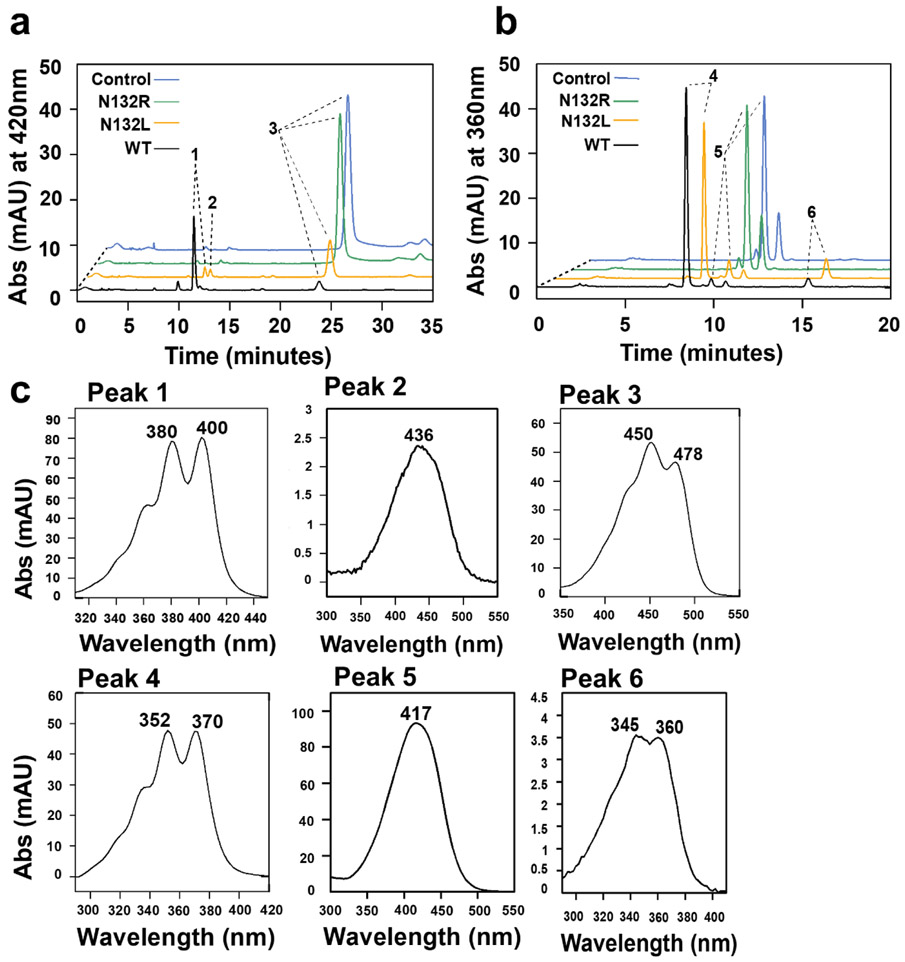
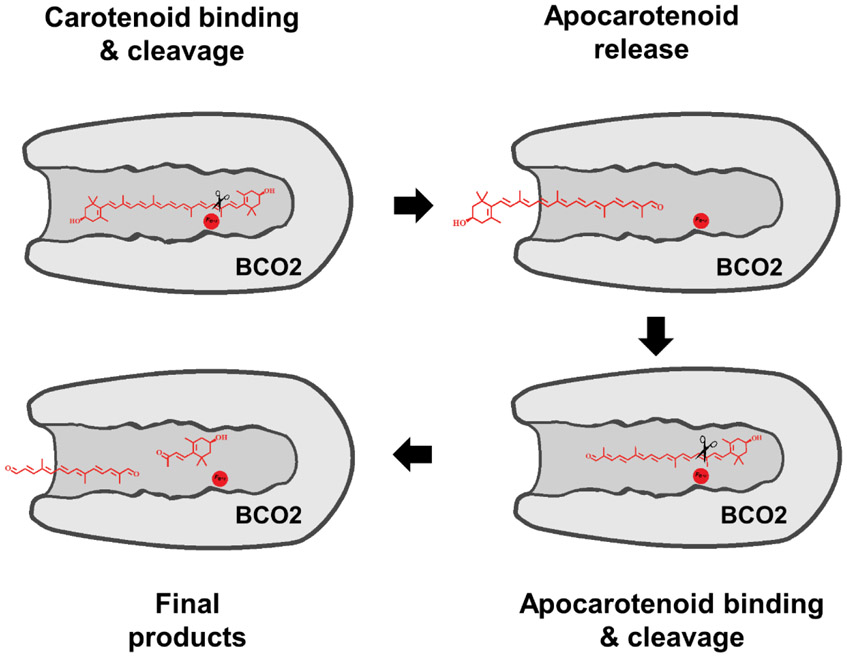
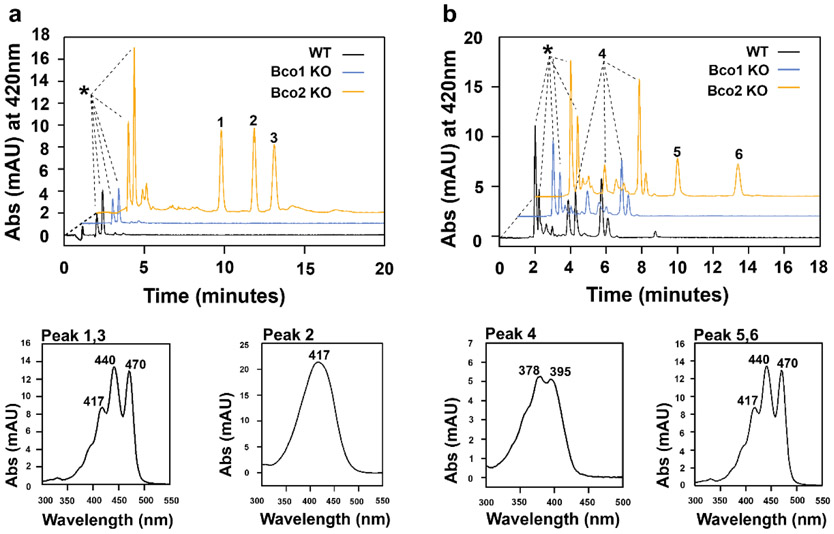
In mammals, carotenoids are converted by two carotenoid cleavage oxygenases into apocarotenoids, including vitamin A. Although knowledge about β-carotene oxygenase-1 (BCO1) and vitamin A metabolism has tremendously increased, the function of β-carotene oxygenase-2 (BCO2) remains less well-defined. We here studied the role of BCO2 in the metabolism of long chain β-apocarotenoids, which recently emerged as putative regulatory molecules in mammalian biology. We showed that recombinant murine BCO2 converted the alcohol, aldehyde, and carboxylic acid of a β-apocarotenoid substrate by oxidative cleavage at position C9,C10 into a β-ionone and a diapocarotenoid product. Chain length variation (C20 to C40) and ionone ring site modifications of the apocarotenoid substrate did not impede catalytic activity or alter the regioselectivity of the double bond cleavage by BCO2. Isotope labeling experiments revealed that the double bond cleavage of an apocarotenoid followed a dioxygenase reaction mechanism. Structural modeling and site directed mutagenesis identified amino acid residues in the substrate tunnel of BCO2 that are critical for apocarotenoid binding and catalytic processing. Mice deficient for BCO2 accumulated apocarotenoids in their livers, indicating that the enzyme engages in apocarotenoid metabolism. Together, our study provides novel structural and functional insights into BCO2 catalysis and establishes the enzyme as a key component of apocarotenoid homeostasis in mice.
Conflict of interest statement
Conflict of interest
The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
