

Review
doi: 10.1084/jem.20201606.
Lipid metabolism and cancer
Affiliations
- PMID: 33601415
- PMCID: PMC7754673
- DOI: 10.1084/jem.20201606
Item in Clipboard
Review
Lipid metabolism and cancer
J Exp Med.
.
Abstract
Dysregulation in lipid metabolism is among the most prominent metabolic alterations in cancer. Cancer cells harness lipid metabolism to obtain energy, components for biological membranes, and signaling molecules needed for proliferation, survival, invasion, metastasis, and response to the tumor microenvironment impact and cancer therapy. Here, we summarize and discuss current knowledge about the advances made in understanding the regulation of lipid metabolism in cancer cells and introduce different approaches that have been clinically used to disrupt lipid metabolism in cancer therapy.
© 2020 Bian et al.
Conflict of interest statement
Disclosures: The authors declare no competing interests exist.
Figures
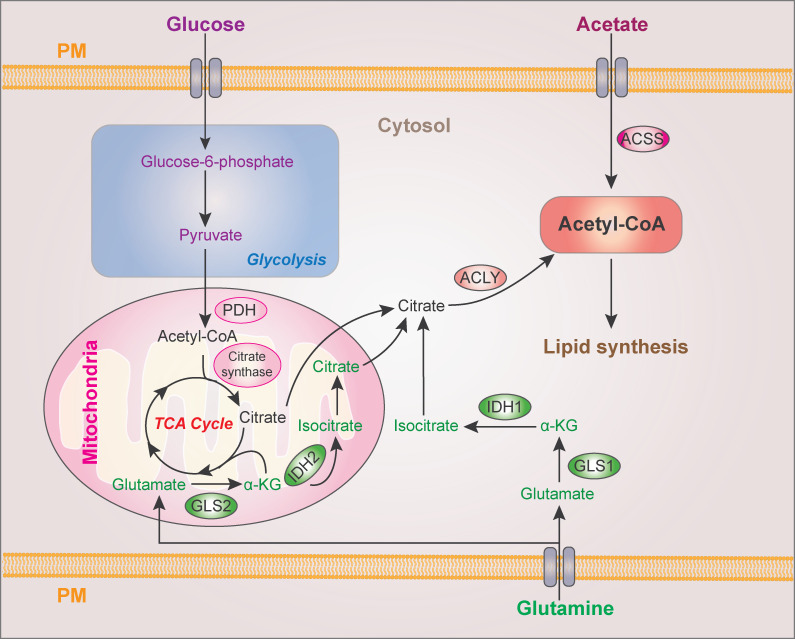
Cytoplasmic acetyl-CoA production for lipid synthesis. Cytoplasmic acetyl-CoA is produced from ACLY-catalyzed citrate and ACSS-catalyzed acetate. Glucose and glutamine contribute to citrate production from mitochondrial pyruvate oxidation in the TCA cycle and reductive carboxylation, respectively. PDH, pyruvate dehydrogenase; α-KG, α-ketoglutarate; GLS, glutaminase; PM, plasma membrane.
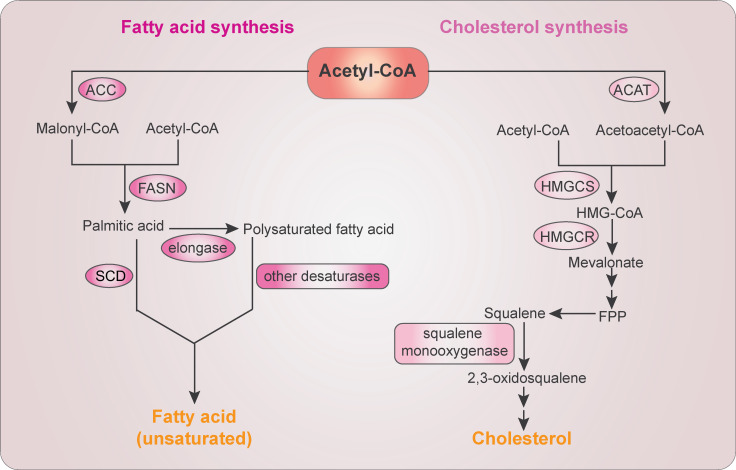
The FA and cholesterol biosynthesis. FA biosynthesis starts with conversion of acetyl-CoA to malonyl-CoA by ACC. Acetyl-CoA and malonyl-CoA are then catalyzed into palmitic acid by FASN. Further elongation is mediated by elongases to form polysaturated FA. Palmitic acid and polysaturated FAs are desaturated into unsaturated FAs by SCD and other fatty acyl-CoA desaturases, respectively. Cholesterol biosynthesis starts with the condensation of two molecules of acetyl-CoA by ACAT to form acetoacetyl-CoA, which is further condensed with a third molecule of acetyl-CoA by HMG-CoA synthase (HMGCS) to form HMG-CoA. HMGCR then reduces HMG-CoA to mevalonate, which is converted to farnesyl pyrophosphate (FPP). Farnesyl pyrophosphate–converted squalene is oxidized by SM to produce 2,3-oxidosqualene, a precursor of cholesterol and sterols.
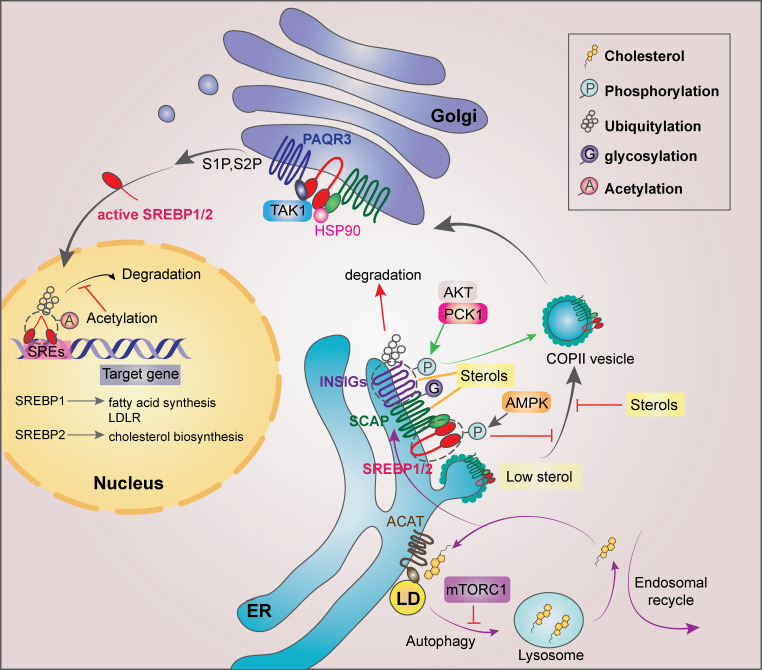
Regulation of SREBP1/2 in cancer cells. SREBP activity can be regulated at multiple levels and at different subcellular localizations. In the ER, sterols bind to SCAP and disrupt the direct interaction between SCAP and COPII for the SREBP ER exit. When sterol level decreases, SCAP dissociates from INSIGs and facilitates the incorporation of SCAP/SREBP into COPII-coated vesicles. mTORC1 suppresses autophagy and subsequent cholesterol trafficking from the lysosome to the ER, leading to SREBP2 activation. Long-chain unsaturated FAs inhibit SREBP activation through inhibition of ubiquitylation of INSIG1. AKT-phosphorylated PCK1 phosphorylates INSIG1/2 and disrupts the bindings of oxysterols to INSIG1/2 for SREBP1/2 activation. In addition, activated AMPK can phosphorylate SREBP1/2 for their retention in the ER. EGFR activation enhances N-glycosylation of SCAP, triggering its dissociation from INSIG1. In the Golgi, SREBP1/2 are cleaved by S1P and S2P, releasing the transcriptionally active SREBP1/2. HSP90 facilitates the SREBP–SCAP complex transit from the ER to the Golgi. PAQR3 potentiates SREBP processing in the Golgi, whereas TAK1-mediated phosphorylation of SREBP1/2 inhibits SREBP. In the nucleus, truncated SREBP1/2 bind to SREs within the promoters of their target genes. GSK3-phosphorylated SREBP1/2 undergoes ubiquitylation and degradation, which can be counteracted by acetylation of the ubiquitylated Lys residues of SREBP1/2. LD, lipid droplet.
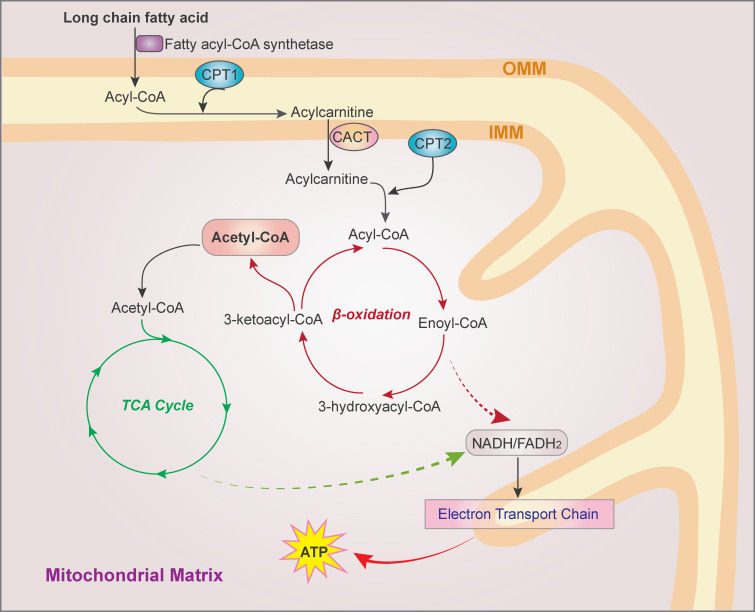
The FAO pathway. FAs are activated to fatty acyl-CoA by fatty acyl-CoA synthetase. On the outer mitochondrial membrane, fatty acyl-CoA is converted to fatty acylcarnitine by CPT1 and shuttled into the mitochondrial matrix. CPT2 on the matrix side of the inner membrane then reconverts acylcarnitine to acyl-CoA, which is cleaved into acetyl-CoA by a repeated four-step cycle catalyzed sequentially by the activity of acyl-CoA dehydrogenase, enoyl-CoA hydratase, 3-hydroxyacyl CoA dehydrogenase, and 3-ketoacyl-CoA thiolase, resulting in the shortening of FAs by two carbons in each cycle. The breakdown product acetyl-CoA enters the TCA cycle, and the produced NADH and FADH2 are coenzymes used in the electron transport chain to generate ATP. CACT, carnitine/acylcarnitine translocase; IMM, inner mitochondrial membrane; OMM, outer mitochondrial membrane.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
