

Differential biosynthesis and cellular permeability explain longitudinal gibberellin gradients in growing roots
- PMID: 33602804
- PMCID: PMC7923382
- DOI: 10.1073/pnas.1921960118
Differential biosynthesis and cellular permeability explain longitudinal gibberellin gradients in growing roots
Abstract
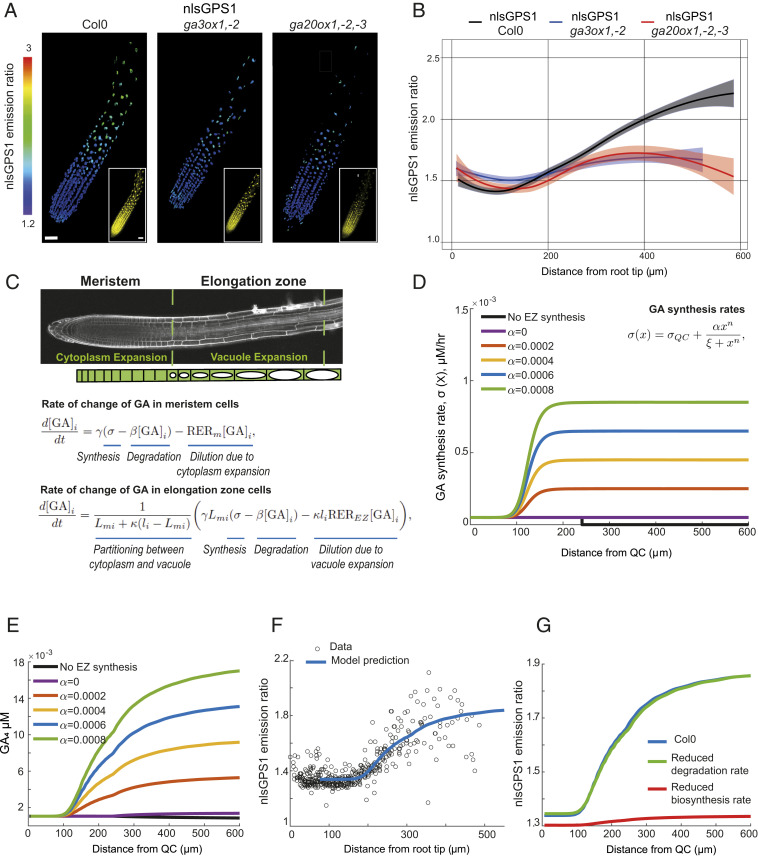
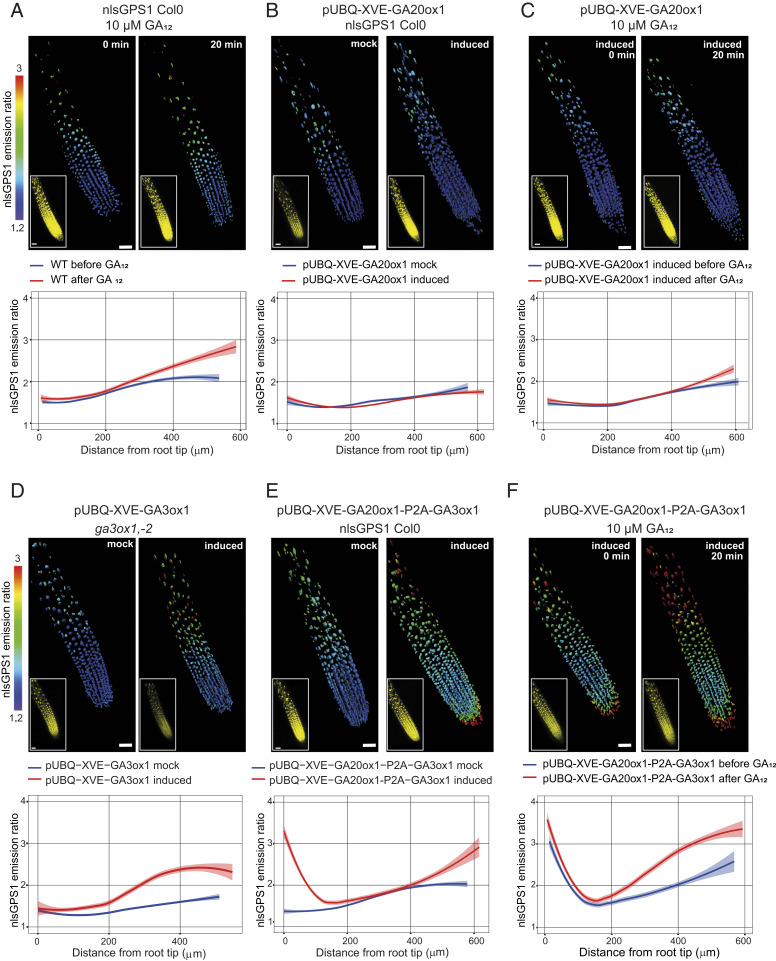
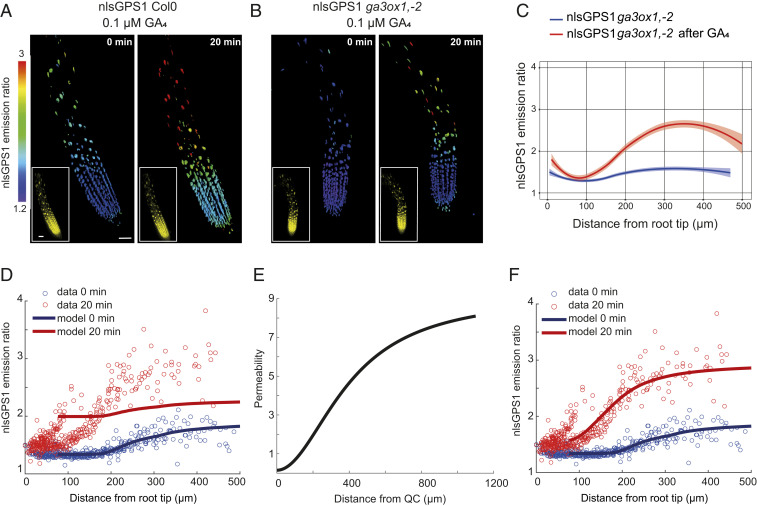
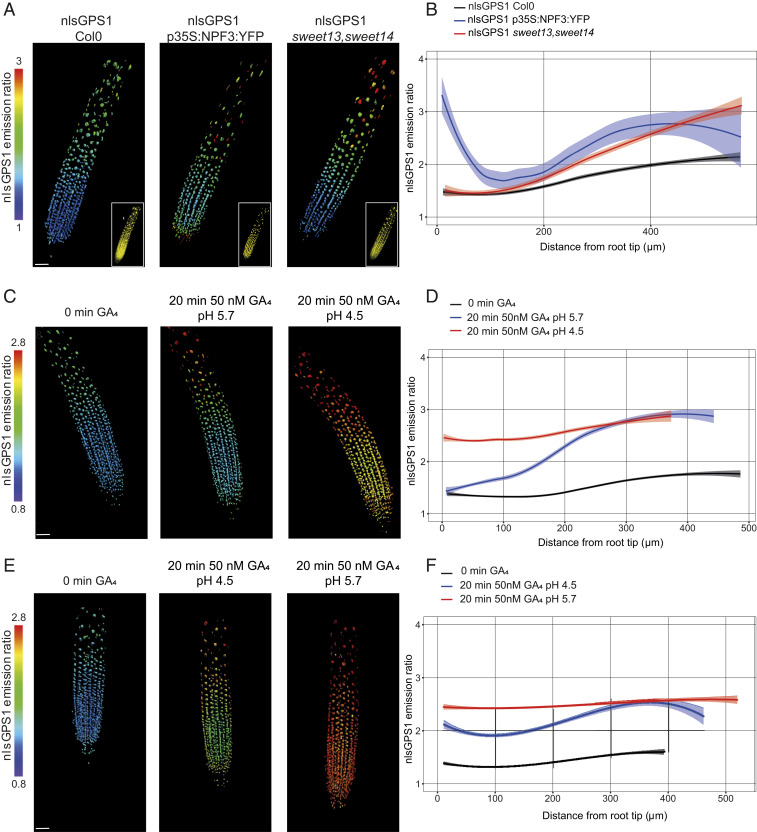
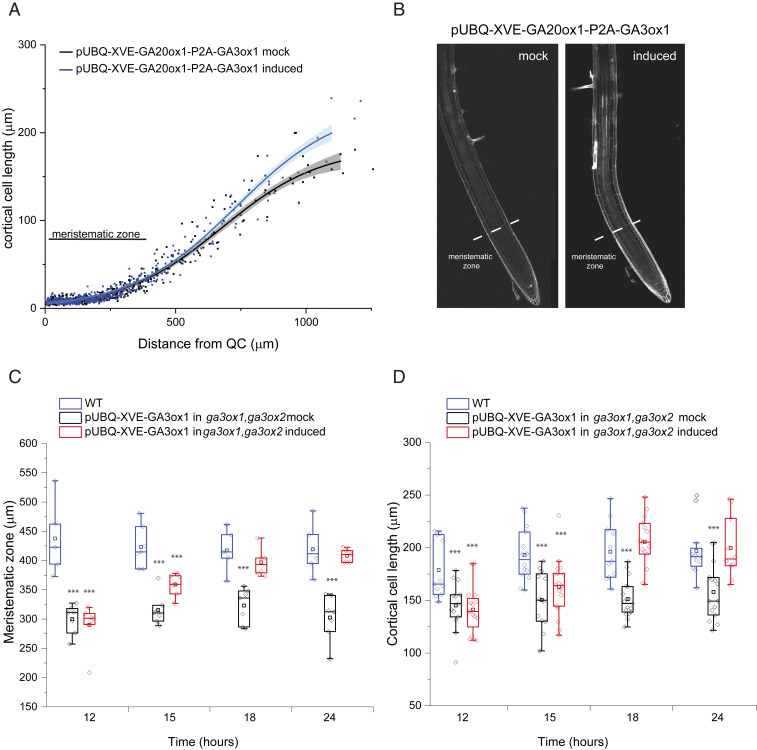
Control over cell growth by mobile regulators underlies much of eukaryotic morphogenesis. In plant roots, cell division and elongation are separated into distinct longitudinal zones and both division and elongation are influenced by the growth regulatory hormone gibberellin (GA). Previously, a multicellular mathematical model predicted a GA maximum at the border of the meristematic and elongation zones. However, GA in roots was recently measured using a genetically encoded fluorescent biosensor, nlsGPS1, and found to be low in the meristematic zone grading to a maximum at the end of the elongation zone. Furthermore, the accumulation rate of exogenous GA was also found to be higher in the elongation zone. It was still unknown which biochemical activities were responsible for these mobile small molecule gradients and whether the spatiotemporal correlation between GA levels and cell length is important for root cell division and elongation patterns. Using a mathematical modeling approach in combination with high-resolution GA measurements in vivo, we now show how differentials in several biosynthetic enzyme steps contribute to the endogenous GA gradient and how differential cellular permeability contributes to an accumulation gradient of exogenous GA. We also analyzed the effects of altered GA distribution in roots and did not find significant phenotypes resulting from increased GA levels or signaling. We did find a substantial temporal delay between complementation of GA distribution and cell division and elongation phenotypes in a GA deficient mutant. Together, our results provide models of how GA gradients are directed and in turn direct root growth.
Keywords: cell growth; gibberellin; hormone biosensor; mathematical modeling; root development.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Hu Y., et al., Gibberellins play an essential role in late embryogenesis of Arabidopsis. Nat. Plants 4, 289–298 (2018). - PubMed
-
- Cheng H., et al., Gibberellin regulates Arabidopsis floral development via suppression of DELLA protein function. Development 131, 1055–1064 (2004). - PubMed
-
- Hedden P., The genes of the green revolution. Trends Genet. 19, 5–9 (2003). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
