

Cell death in head and neck cancer pathogenesis and treatment
- PMID: 33602906
- PMCID: PMC7893032
- DOI: 10.1038/s41419-021-03474-5
Cell death in head and neck cancer pathogenesis and treatment
Abstract
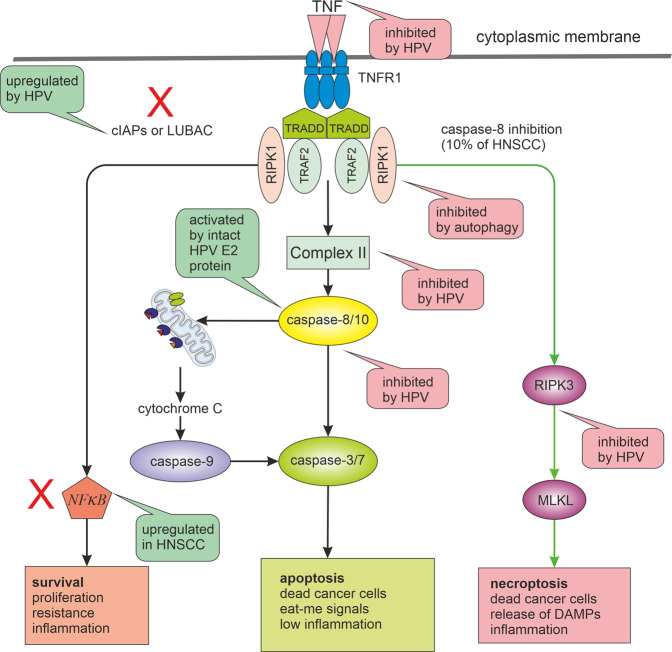
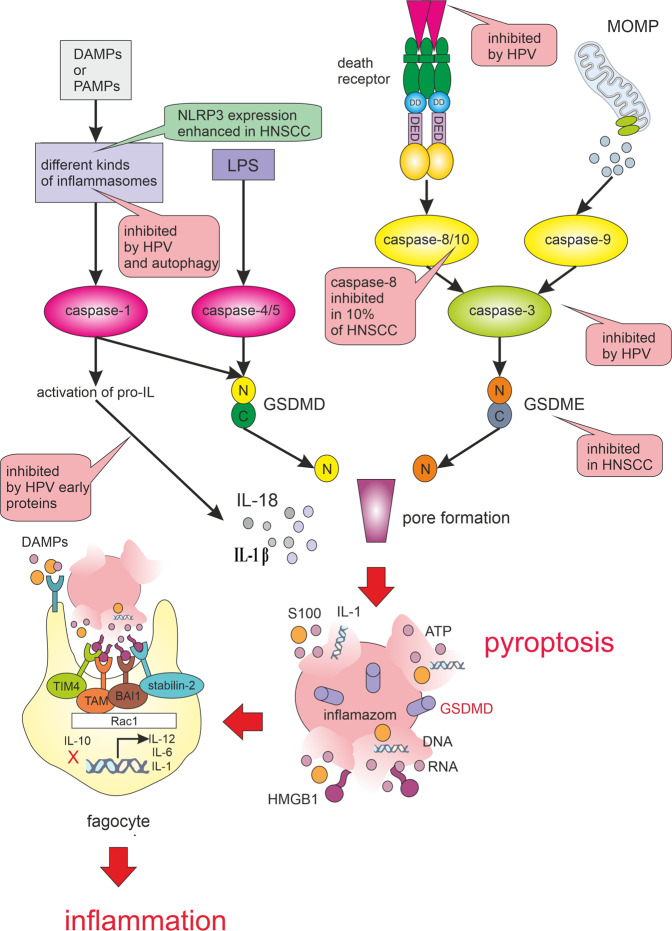
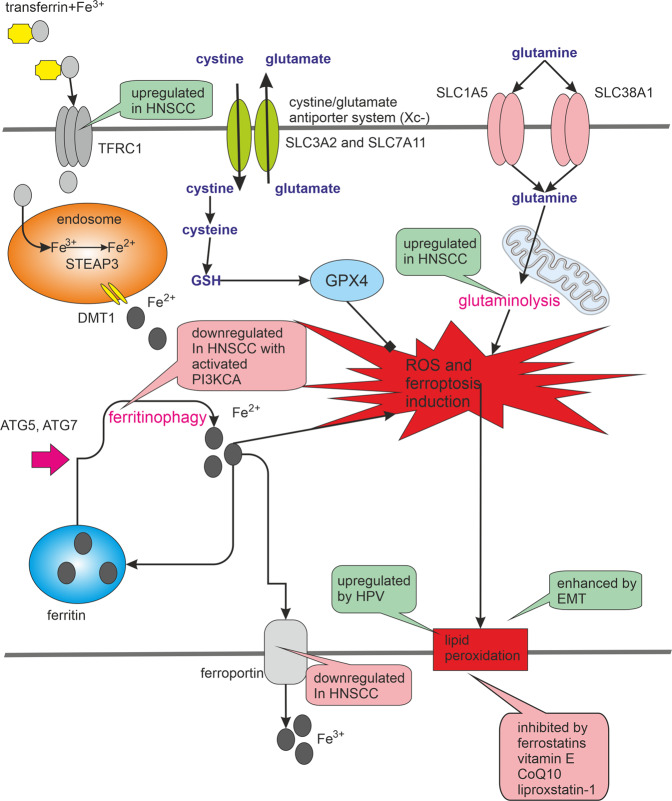
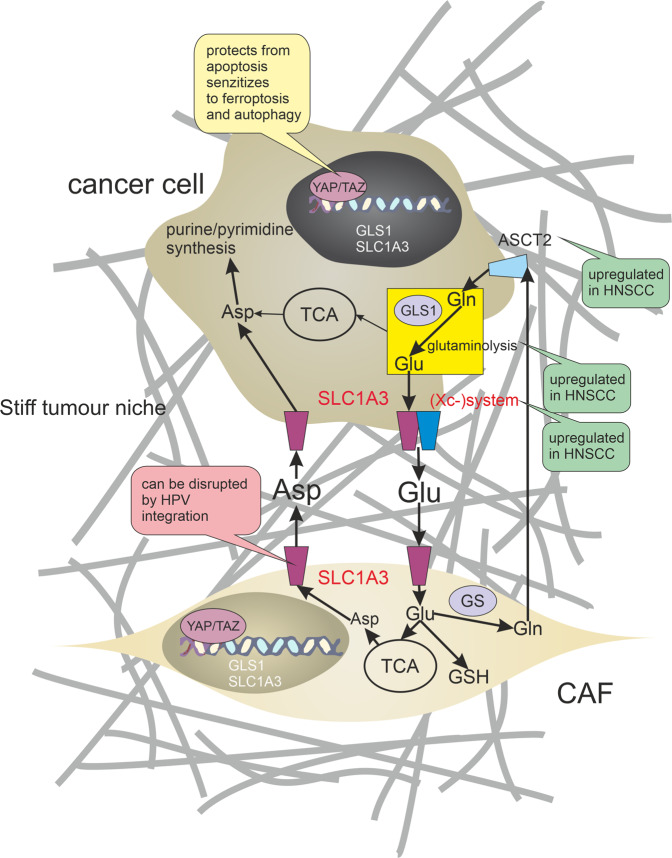
Many cancer therapies aim to trigger apoptosis in cancer cells. Nevertheless, the presence of oncogenic alterations in these cells and distorted composition of tumour microenvironment largely limit the clinical efficacy of this type of therapy. Luckily, scientific consensus describes about 10 different cell death subroutines with different regulatory pathways and cancer cells are probably not able to avoid all of cell death types at once. Therefore, a focused and individualised therapy is needed to address the specific advantages and disadvantages of individual tumours. Although much is known about apoptosis, therapeutic opportunities of other cell death pathways are often neglected. Molecular heterogeneity of head and neck squamous cell carcinomas (HNSCC) causing unpredictability of the clinical response represents a grave challenge for oncologists and seems to be a critical component of treatment response. The large proportion of this clinical heterogeneity probably lies in alterations of cell death pathways. How exactly cells die is very important because the predominant type of cell death can have multiple impacts on the therapeutic response as cell death itself acts as a second messenger. In this review, we discuss the different types of programmed cell death (PCD), their connection with HNSCC pathogenesis and possible therapeutic windows that result from specific sensitivity to some form of PCD in some clinically relevant subgroups of HNSCC.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
