

Effect of palmitoylation on the dimer formation of the human dopamine transporter
- PMID: 33602981
- PMCID: PMC7893171
- DOI: 10.1038/s41598-021-83374-y
Effect of palmitoylation on the dimer formation of the human dopamine transporter
Abstract
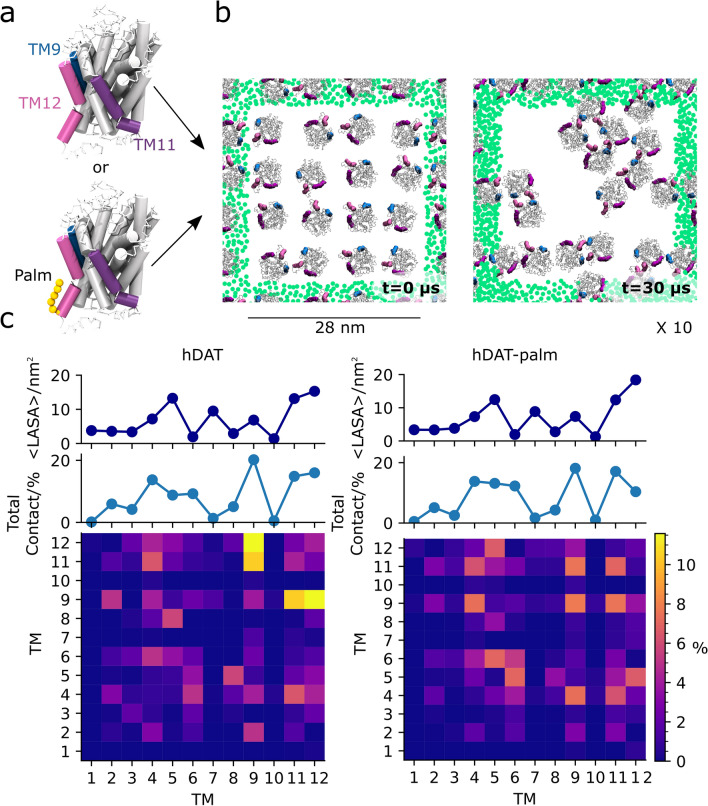
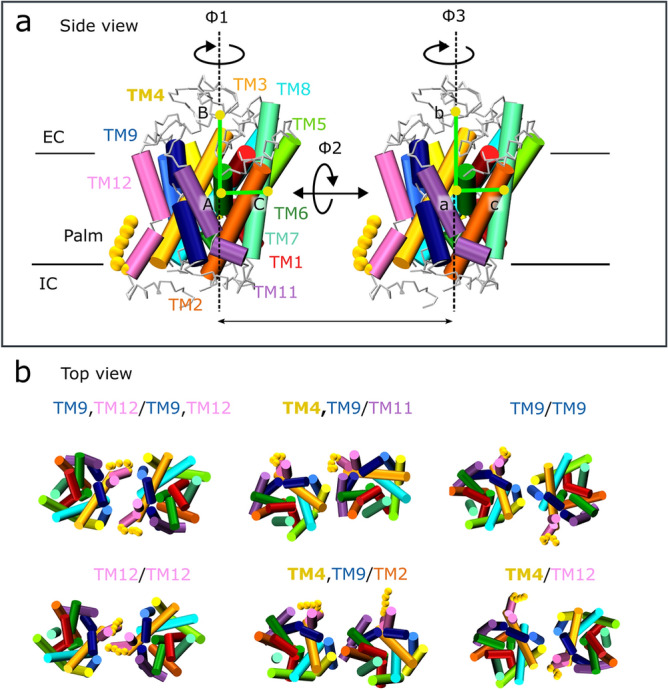
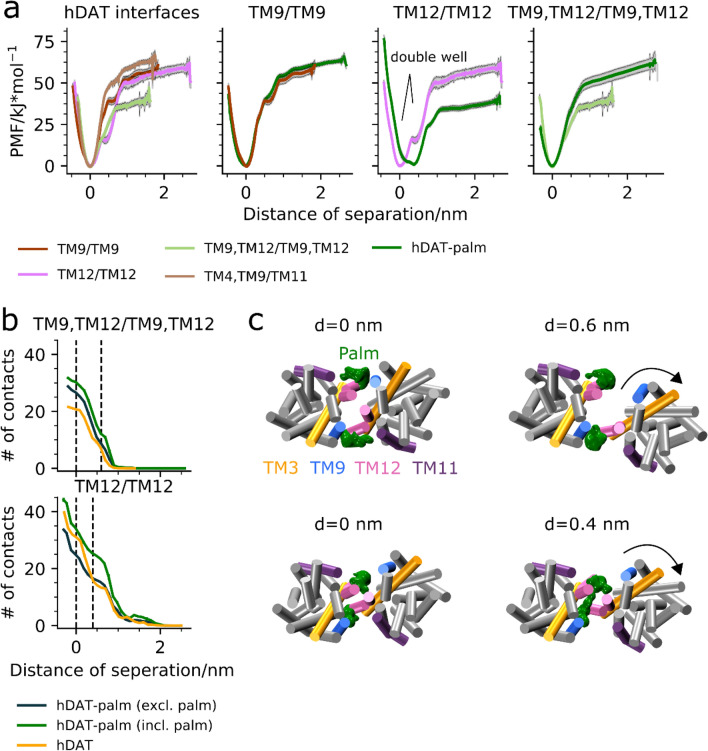
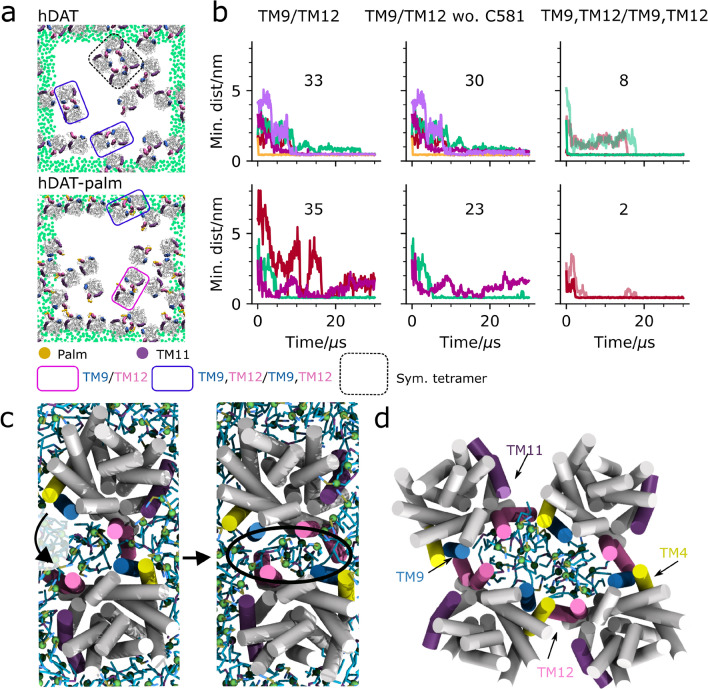
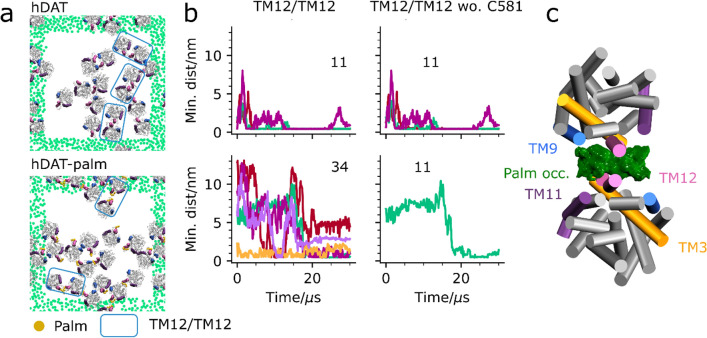
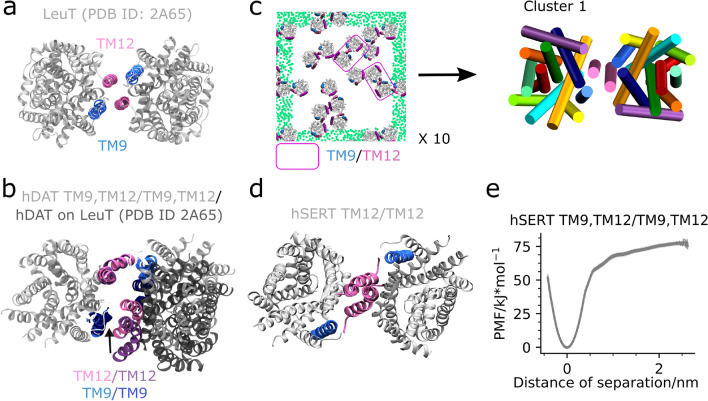
The human dopamine transporter (hDAT) is one in three members of the monoamine transporter family (MAT). hDAT is essential for regulating the dopamine concentration in the synaptic cleft through dopamine reuptake into the presynaptic neuron; thereby controlling hDAT dopamine signaling. Dysfunction of the transporter is linked to several psychiatric disorders. hDAT and the other MATs have been shown to form oligomers in the plasma membrane, but only limited data exists on which dimeric and higher order oligomeric states are accessible and energetically favorable. In this work, we present several probable dimer conformations using computational coarse-grained self-assembly simulations and assess the relative stability of the different dimer conformations using umbrella sampling replica exchange molecular dynamics. Overall, the dimer conformations primarily involve TM9 and/or TM11 and/or TM12 at the interface. Furthermore, we show that a palmitoyl group (palm) attached to hDAT on TM12 modifies the free energy of separation for interfaces involving TM12, suggesting that S-palmitoylation may change the relative abundance of dimers involving TM12 in a biological context. Finally, a comparison of the identified interfaces of hDAT and palmitoylated hDAT to the human serotonin transporter interfaces and the leucine transporter interface, suggests similar dimer conformations across these protein family.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Norregaard L, Gether U. The monoamine neurotransmitter transporters: structure, conformational changes and molecular gating. Curr. Opin. Drug. Disc. Dev. 2001;4:591–601. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
