

Discovery of a novel lactate dehydrogenase tetramerization domain using epitope mapping and peptides
- PMID: 33607109
- PMCID: PMC8010463
- DOI: 10.1016/j.jbc.2021.100422
Discovery of a novel lactate dehydrogenase tetramerization domain using epitope mapping and peptides
Abstract
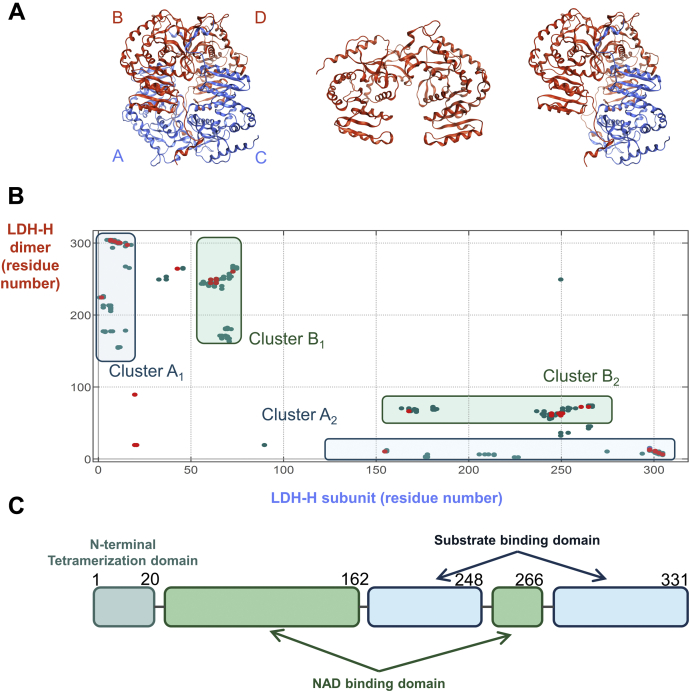
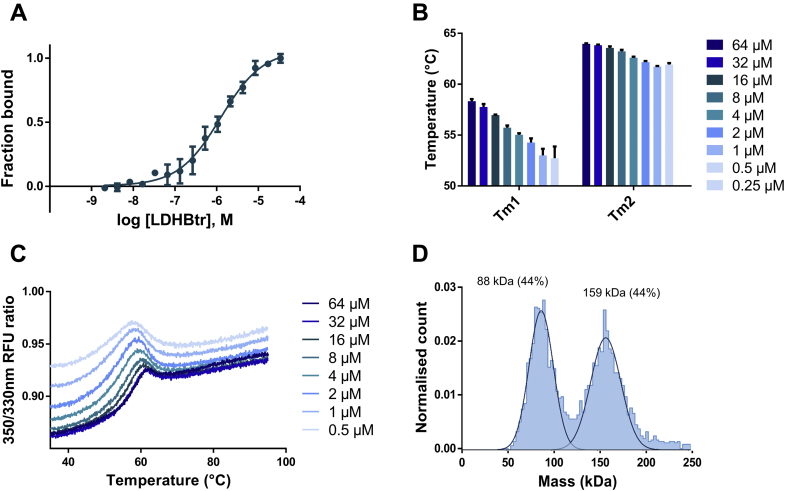
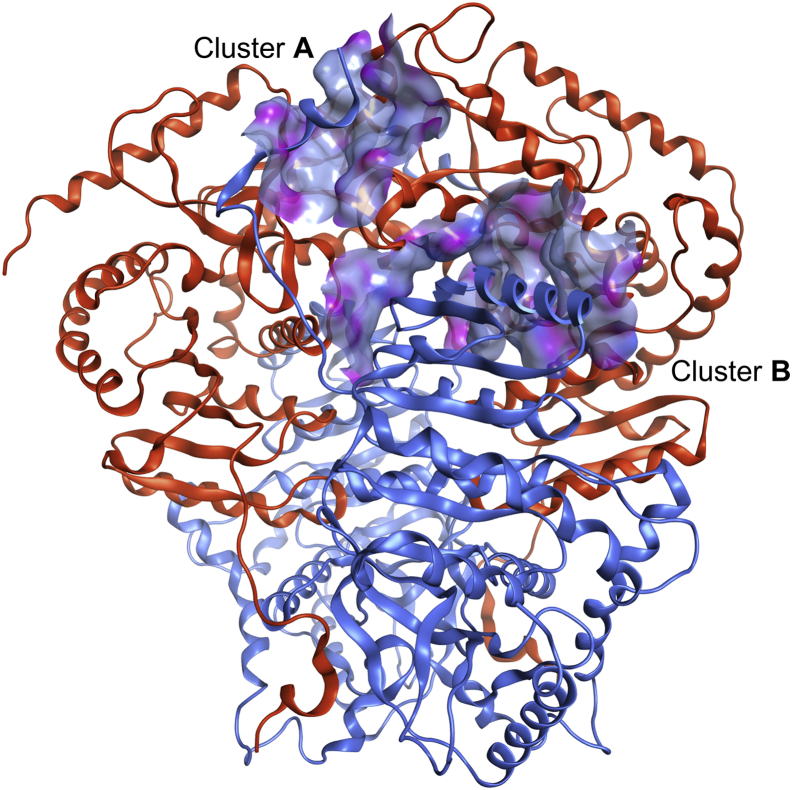
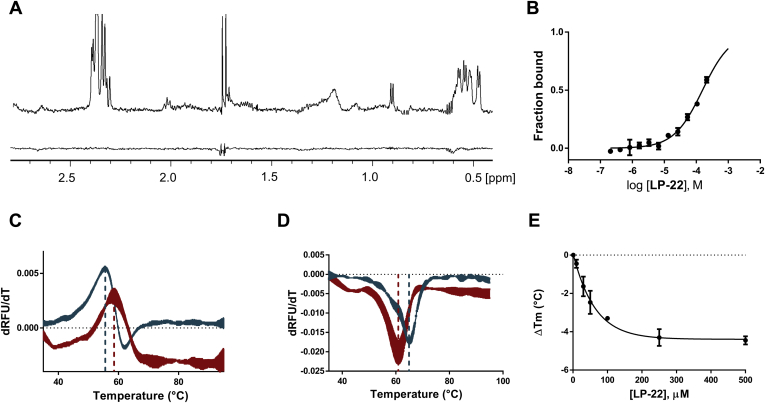
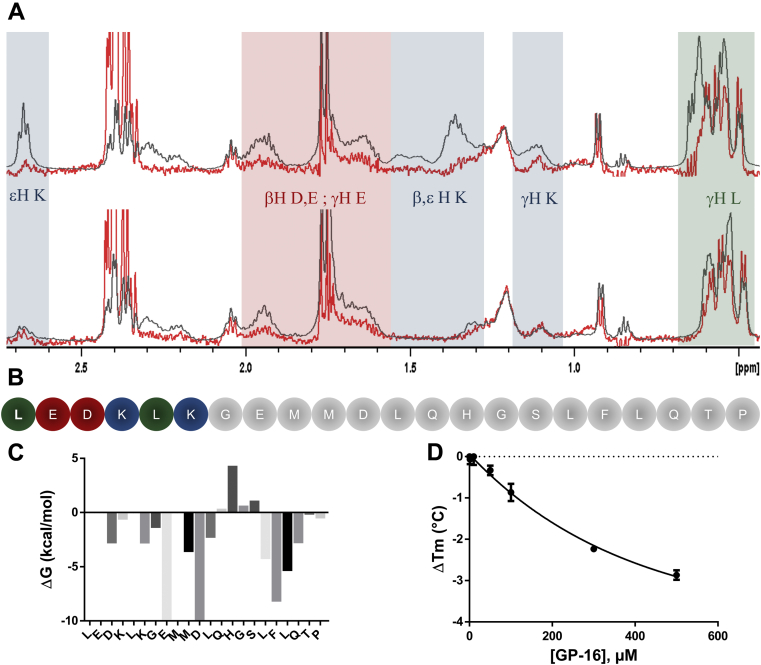
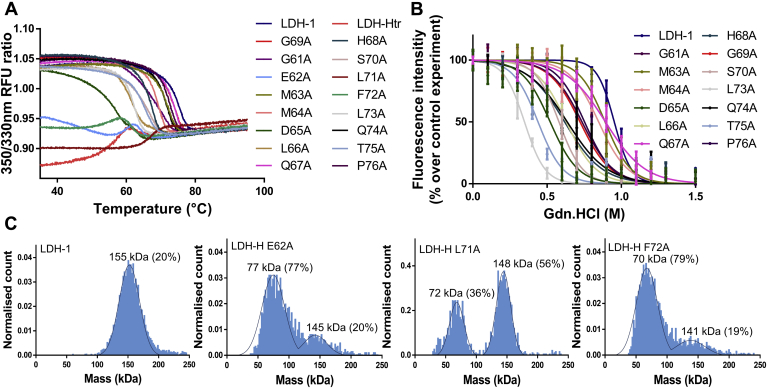
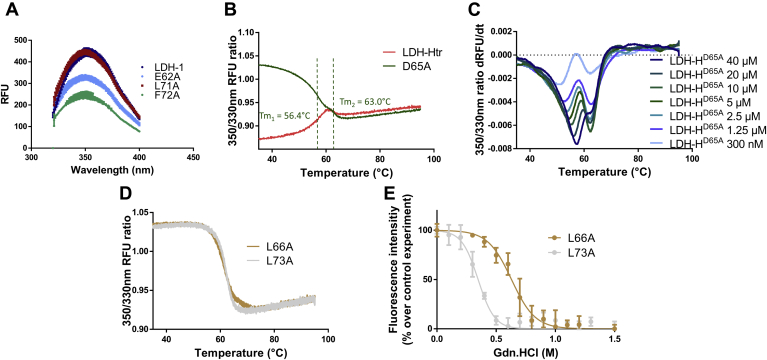
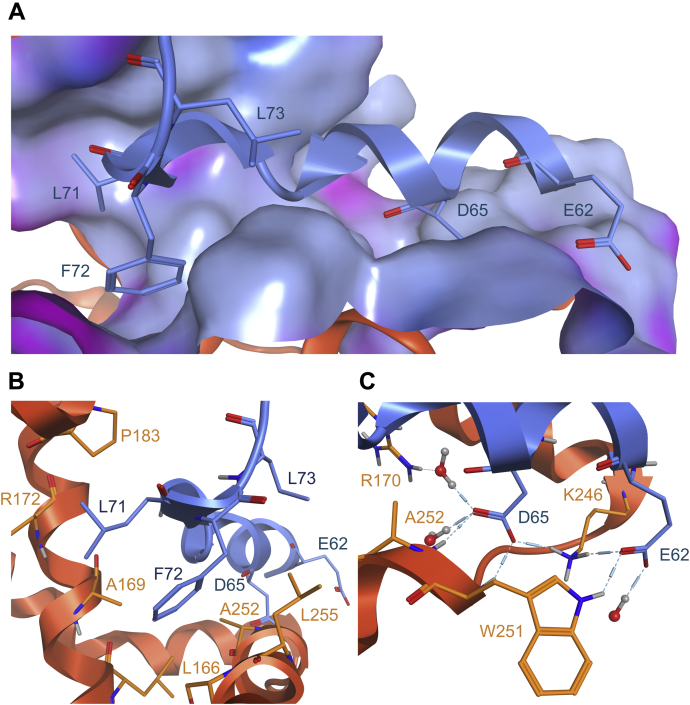
Despite being initially regarded as a metabolic waste product, lactate is now considered to serve as a primary fuel for the tricarboxylic acid cycle in cancer cells. At the core of lactate metabolism, lactate dehydrogenases (LDHs) catalyze the interconversion of lactate to pyruvate and as such represent promising targets in cancer therapy. However, direct inhibition of the LDH active site is challenging from physicochemical and selectivity standpoints. However, LDHs are obligate tetramers. Thus, targeting the LDH tetrameric interface has emerged as an appealing strategy. In this work, we examine a dimeric construct of truncated human LDH to search for new druggable sites. We report the identification and characterization of a new cluster of interactions in the LDH tetrameric interface. Using nanoscale differential scanning fluorimetry, chemical denaturation, and mass photometry, we identified several residues (E62, D65, L71, and F72) essential for LDH tetrameric stability. Moreover, we report a family of peptide ligands based on this cluster of interactions. We next demonstrated these ligands to destabilize tetrameric LDHs through binding to this new tetrameric interface using nanoscale differential scanning fluorimetry, NMR water-ligand observed via gradient spectroscopy, and microscale thermophoresis. Altogether, this work provides new insights on the LDH tetrameric interface as well as valuable pharmacological tools for the development of LDH tetramer disruptors.
Keywords: NMR; WaterLOGSY; biophysics; cancer; disruptors; lactate dehydrogenases; mass photometry; microscale thermophoresis; nanoDSF; oligomerization; protein–protein interaction.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare the following competing financial interest(s): L. T., L. B., P. S., and R. F. are inventors of European Patent Application EP19172347.7, LDH inhibitor polypeptides for use in treatment of cancer; L. T., M. L., P. S., and R. F. are inventors of European Patent Application EP21154636.1, polypeptide inhibitors of LDH activity for use in cancer therapy. The authors declare no other conflicts of interest.
Figures
References
-
- Beckert S., Farrahi F., Aslam R.S., Scheuenstuhl H., Königsrainer A., Hussain M.Z., Hunt T.K. Lactate stimulates endothelial cell migration. Wound Repair Regen. 2006;14:321–324. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
