

Mitochondrial DNA editing in mice with DddA-TALE fusion deaminases
- PMID: 33608520
- PMCID: PMC7895935
- DOI: 10.1038/s41467-021-21464-1
Mitochondrial DNA editing in mice with DddA-TALE fusion deaminases
Abstract
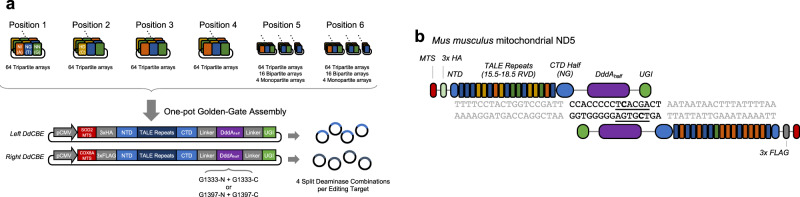
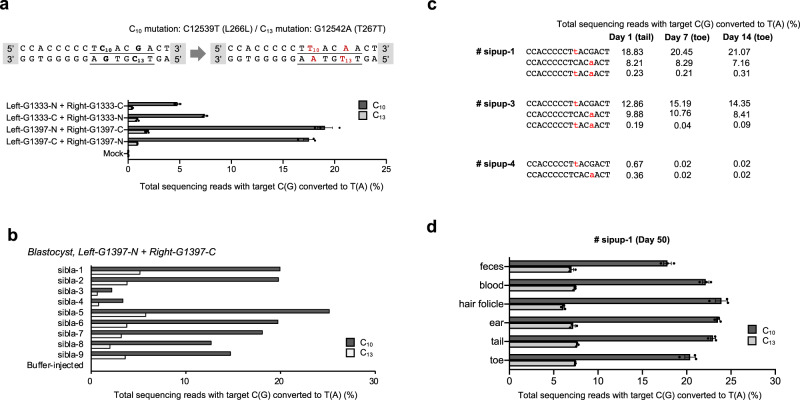
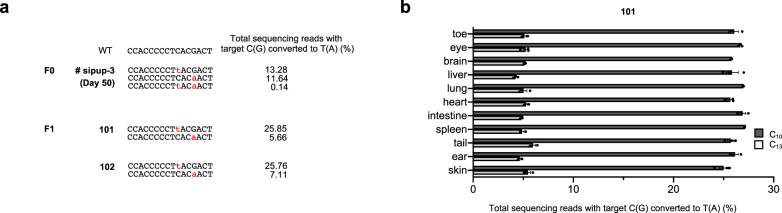
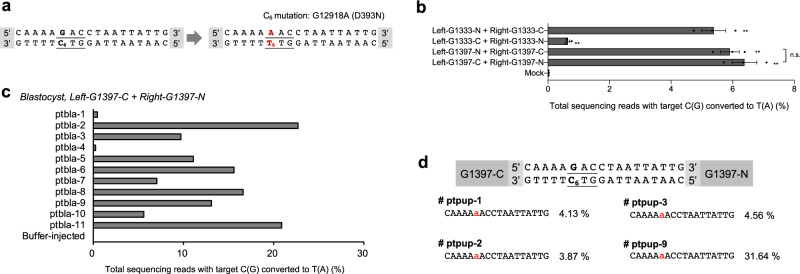
DddA-derived cytosine base editors (DdCBEs), composed of the split interbacterial toxin DddAtox, transcription activator-like effector (TALE), and uracil glycosylase inhibitor (UGI), enable targeted C-to-T base conversions in mitochondrial DNA (mtDNA). Here, we demonstrate highly efficient mtDNA editing in mouse embryos using custom-designed DdCBEs. We target the mitochondrial gene, MT-ND5 (ND5), which encodes a subunit of NADH dehydrogenase that catalyzes NADH dehydration and electron transfer to ubiquinone, to obtain several mtDNA mutations, including m.G12918A associated with human mitochondrial diseases and m.C12336T that incorporates a premature stop codon, creating mitochondrial disease models in mice and demonstrating a potential for the treatment of mitochondrial disorders.
Conflict of interest statement
J.-S.K. is a cofounder of and holds stock in ToolGen. The other authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
