

Microbiota-Driven Activation of Intrahepatic B Cells Aggravates NASH Through Innate and Adaptive Signaling
- PMID: 33609303
- PMCID: PMC8377092
- DOI: 10.1002/hep.31755
Microbiota-Driven Activation of Intrahepatic B Cells Aggravates NASH Through Innate and Adaptive Signaling
Abstract
Background and aims: Nonalcoholic steatohepatitis is rapidly becoming the leading cause of liver failure and indication for liver transplantation. Hepatic inflammation is a key feature of NASH but the immune pathways involved in this process are poorly understood. B lymphocytes are cells of the adaptive immune system that are critical regulators of immune responses. However, the role of B cells in the pathogenesis of NASH and the potential mechanisms leading to their activation in the liver are unclear.
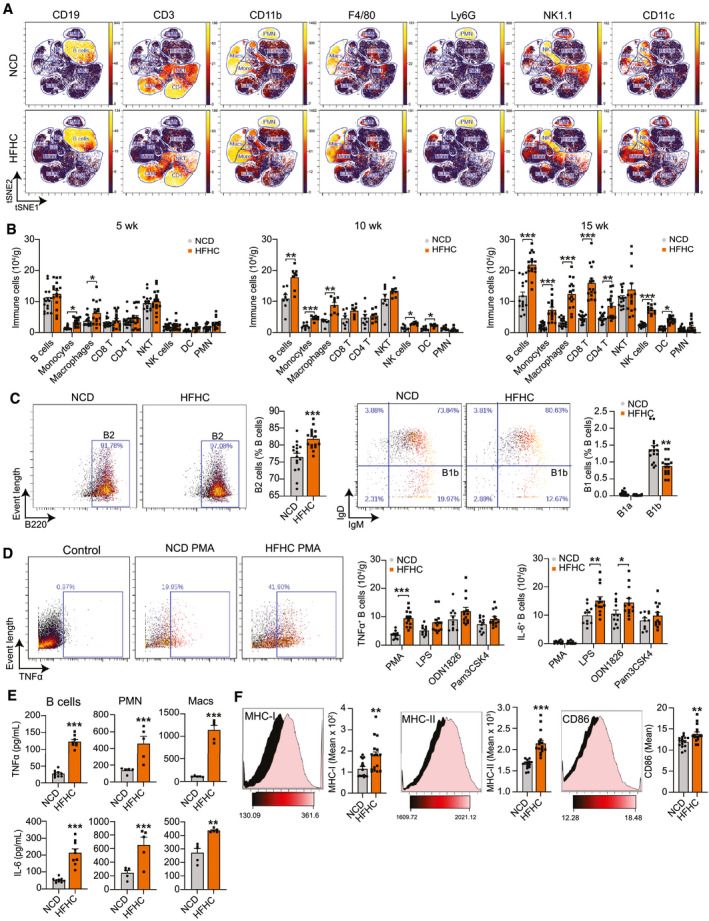
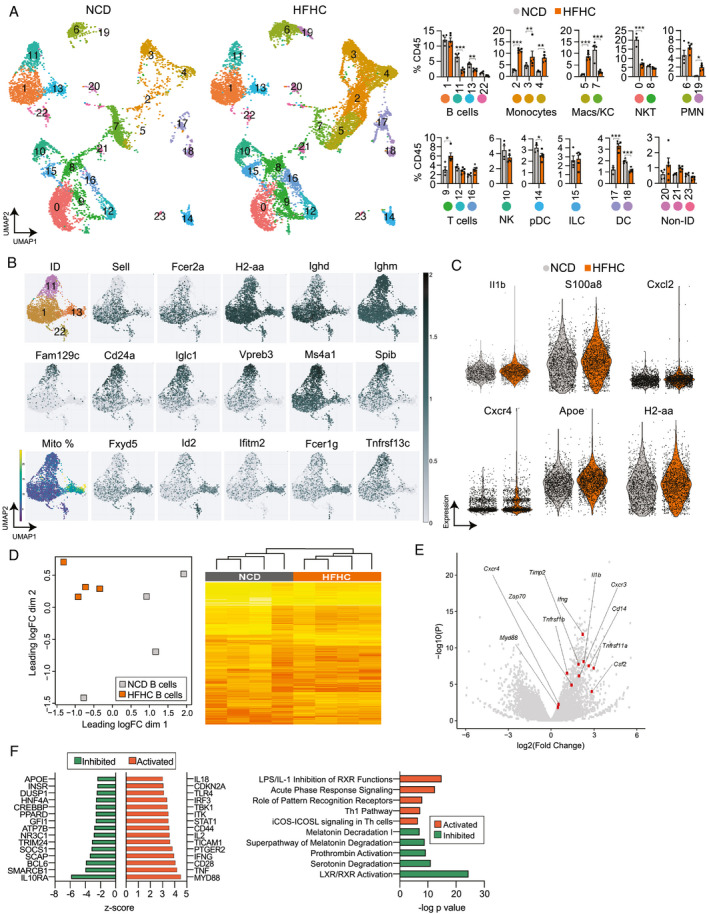
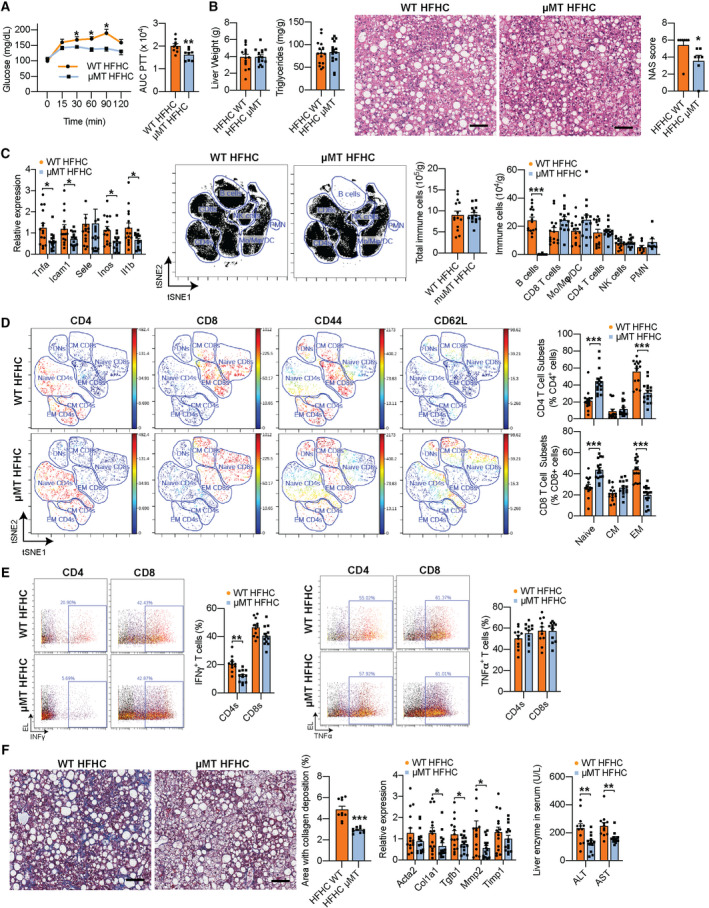
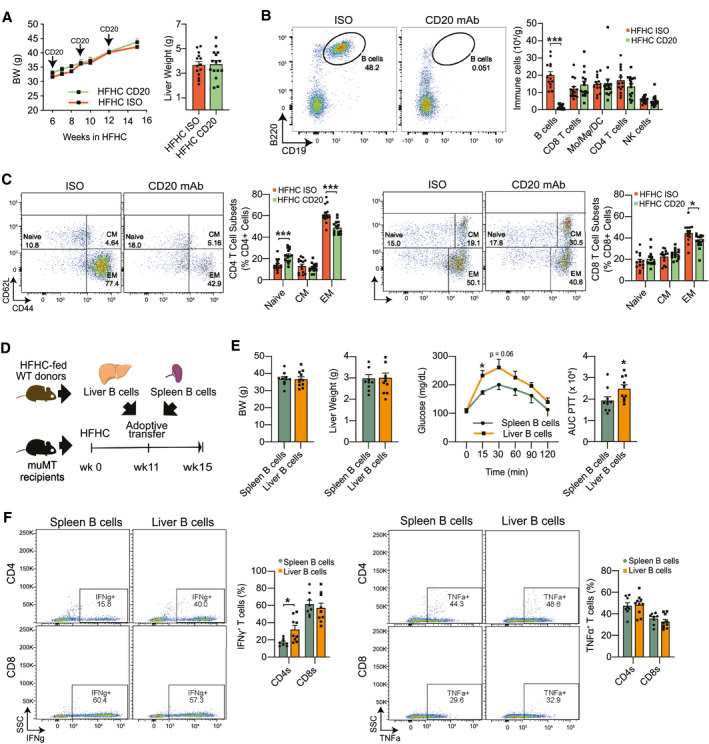
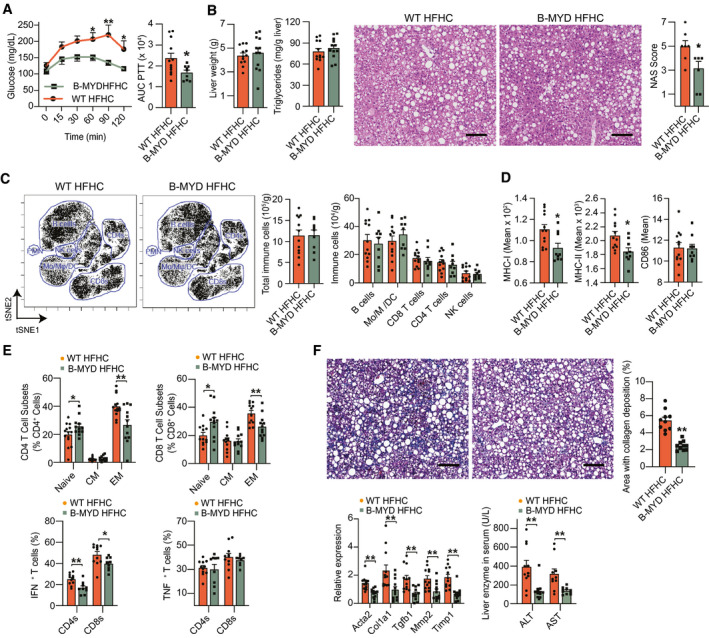
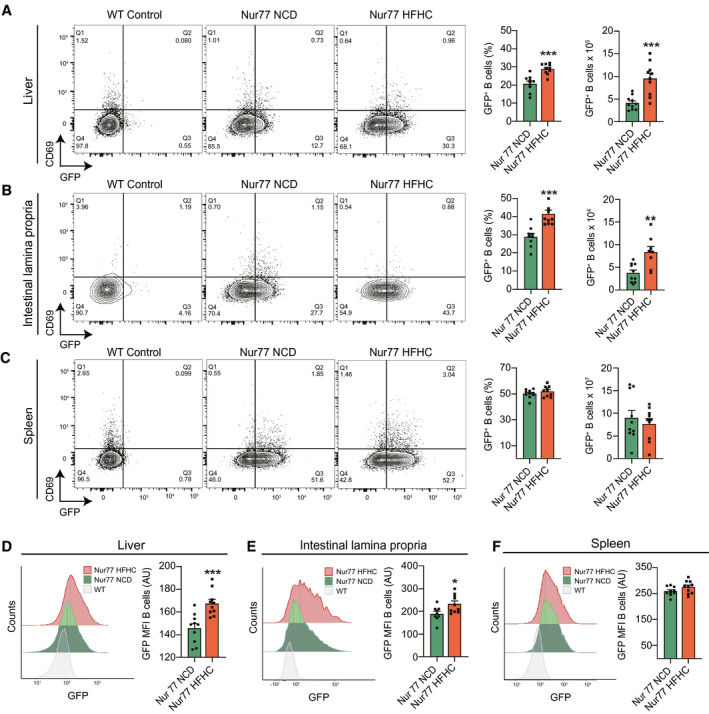
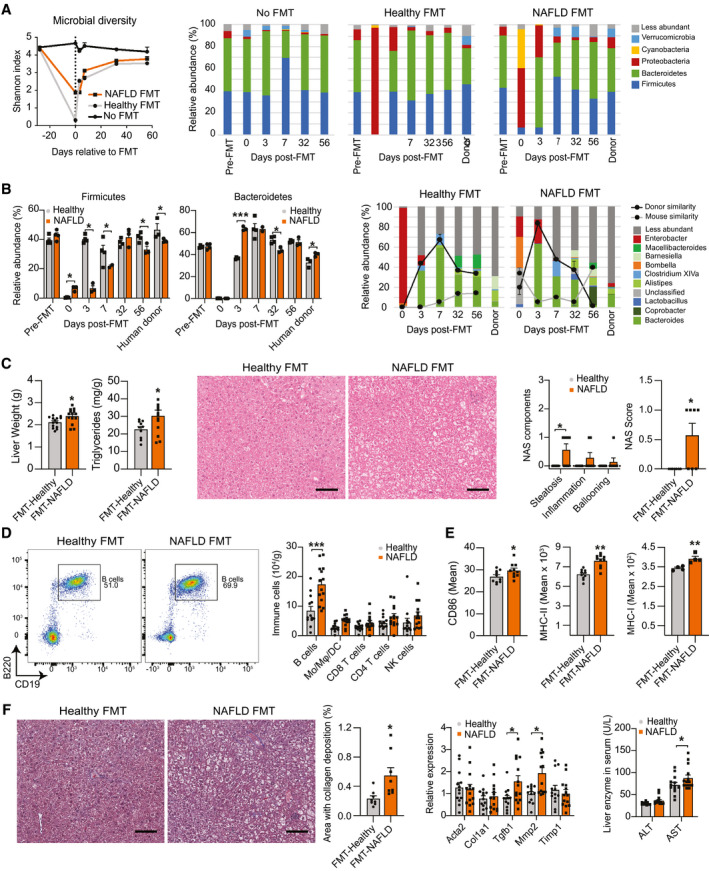
Approach and results: In this study, we report that NASH livers accumulate B cells with elevated pro-inflammatory cytokine secretion and antigen-presentation ability. Single-cell and bulk RNA sequencing of intrahepatic B cells from mice with NASH unveiled a transcriptional landscape that reflects their pro-inflammatory function. Accordingly, B-cell deficiency ameliorated NASH progression, and adoptively transferring B cells from NASH livers recapitulates the disease. Mechanistically, B-cell activation during NASH involves signaling through the innate adaptor myeloid differentiation primary response protein 88 (MyD88) as B cell-specific deletion of MyD88 reduced hepatic T cell-mediated inflammation and fibrosis, but not steatosis. In addition, activation of intrahepatic B cells implicates B cell-receptor signaling, delineating a synergy between innate and adaptive mechanisms of antigen recognition. Furthermore, fecal microbiota transplantation of human NAFLD gut microbiotas into recipient mice promoted the progression of NASH by increasing the accumulation and activation of intrahepatic B cells, suggesting that gut microbial factors drive the pathogenic function of B cells during NASH.
Conclusion: Our findings reveal that a gut microbiota-driven activation of intrahepatic B cells leads to hepatic inflammation and fibrosis during the progression of NASH through innate and adaptive immune mechanisms.
© 2021 The Authors. Hepatology published by Wiley Periodicals LLC on behalf of American Association for the Study of Liver Diseases.
Figures
Comment in
-
The role of B cells in metabolic (dysfunction)-associated fatty liver disease.Hepatobiliary Surg Nutr. 2021 Dec;10(6):875-877. doi: 10.21037/hbsn-21-404. Hepatobiliary Surg Nutr. 2021. PMID: 35004959 Free PMC article. No abstract available.
References
-
- Younossi Z, Anstee QM, Marietti M, Hardy T, Henry L, Eslam M, et al. Global burden of NAFLD and NASH: trends, predictions, risk factors and prevention. Nat Rev Gastroenterol Hepatol 2018;15:11‐20. - PubMed
-
- Tilg H, Moschen AR. Evolution of inflammation in nonalcoholic fatty liver disease: the multiple parallel hits hypothesis. Hepatology 2010;52:1836‐1846. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
