

Role of NAD+ in regulating cellular and metabolic signaling pathways
- PMID: 33609766
- PMCID: PMC7973386
- DOI: 10.1016/j.molmet.2021.101195
Role of NAD+ in regulating cellular and metabolic signaling pathways
Abstract
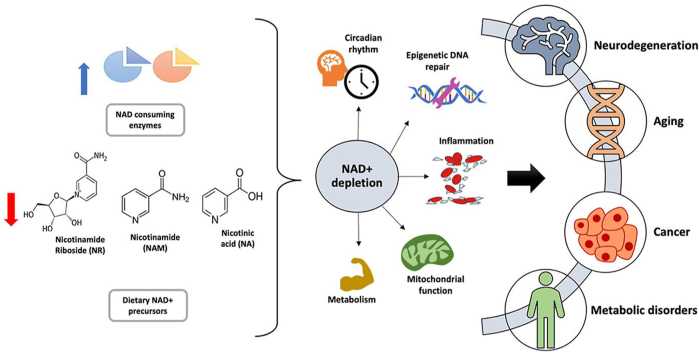
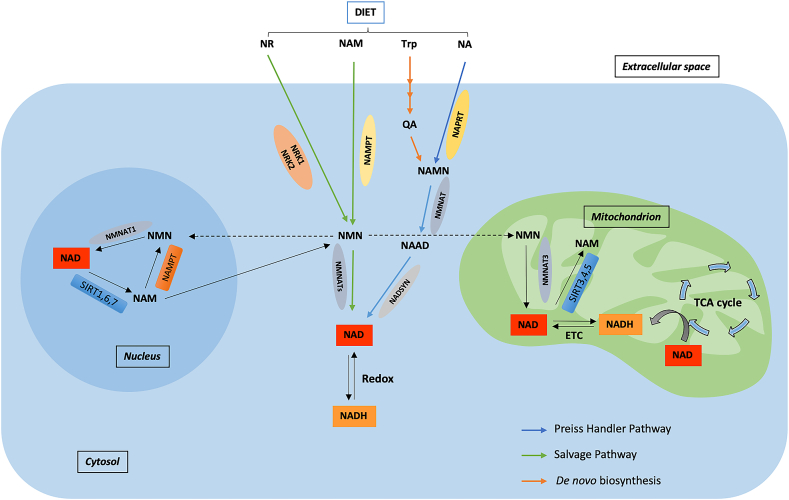
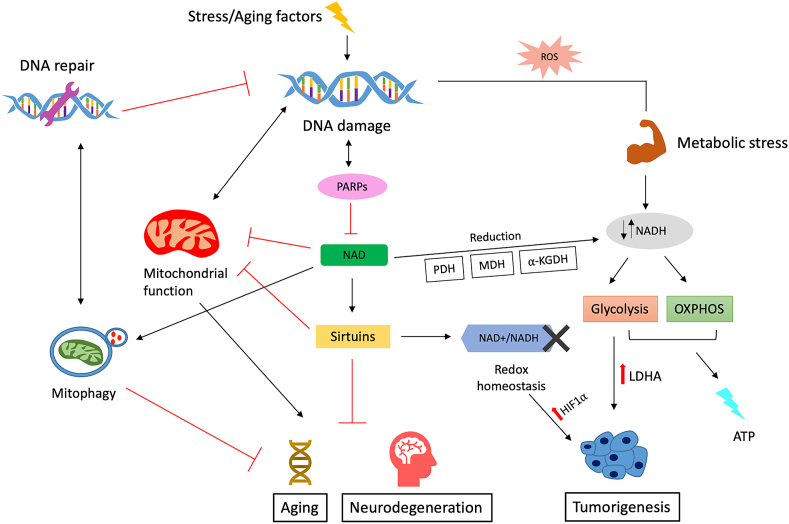
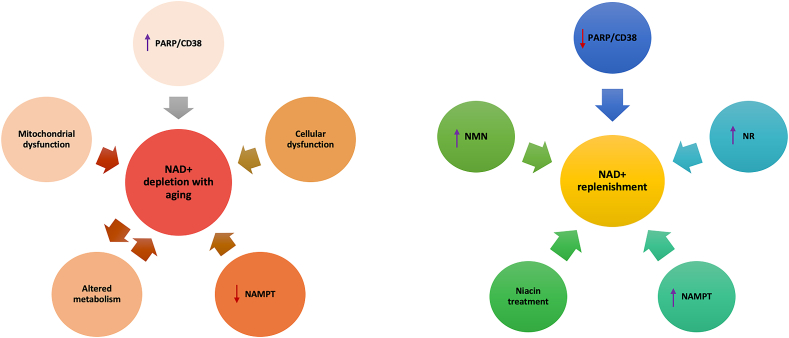
Background: Nicotinamide adenine dinucleotide (NAD+), a critical coenzyme present in every living cell, is involved in a myriad of metabolic processes associated with cellular bioenergetics. For this reason, NAD+ is often studied in the context of aging, cancer, and neurodegenerative and metabolic disorders.
Scope of review: Cellular NAD+ depletion is associated with compromised adaptive cellular stress responses, impaired neuronal plasticity, impaired DNA repair, and cellular senescence. Increasing evidence has shown the efficacy of boosting NAD+ levels using NAD+ precursors in various diseases. This review provides a comprehensive understanding into the role of NAD+ in aging and other pathologies and discusses potential therapeutic targets.
Major conclusions: An alteration in the NAD+/NADH ratio or the NAD+ pool size can lead to derailment of the biological system and contribute to various neurodegenerative disorders, aging, and tumorigenesis. Due to the varied distribution of NAD+/NADH in different locations within cells, the direct role of impaired NAD+-dependent processes in humans remains unestablished. In this regard, longitudinal studies are needed to quantify NAD+ and its related metabolites. Future research should focus on measuring the fluxes through pathways associated with NAD+ synthesis and degradation.
Keywords: Aging; Cancer; Metabolism; NAD(+); Neurodegeneration; Sirtuins.
Copyright © 2021 The Author(s). Published by Elsevier GmbH.. All rights reserved.
Figures
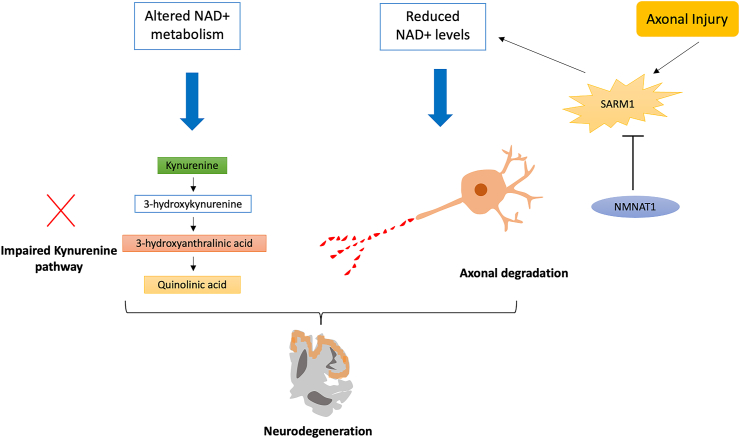
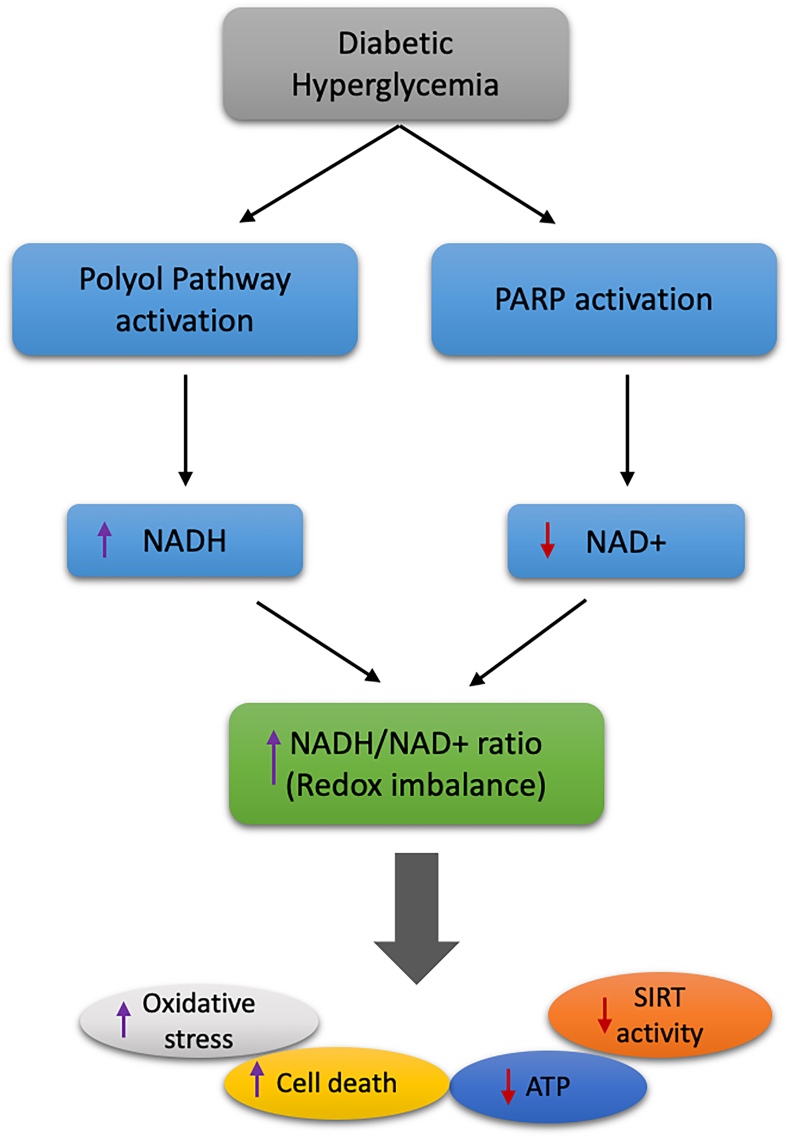
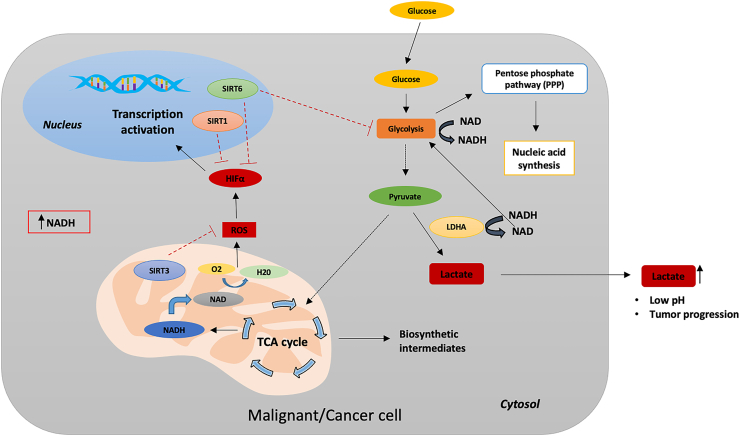
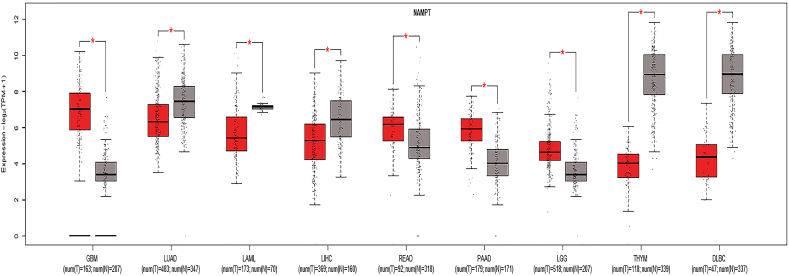
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
