

Modeling Circadian Phototransduction: Retinal Neurophysiology and Neuroanatomy
- PMID: 33613175
- PMCID: PMC7892603
- DOI: 10.3389/fnins.2020.615305
Modeling Circadian Phototransduction: Retinal Neurophysiology and Neuroanatomy
Abstract
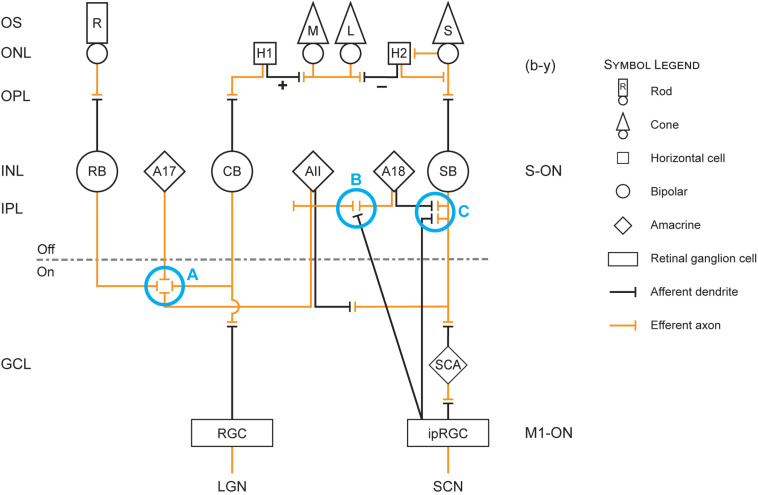
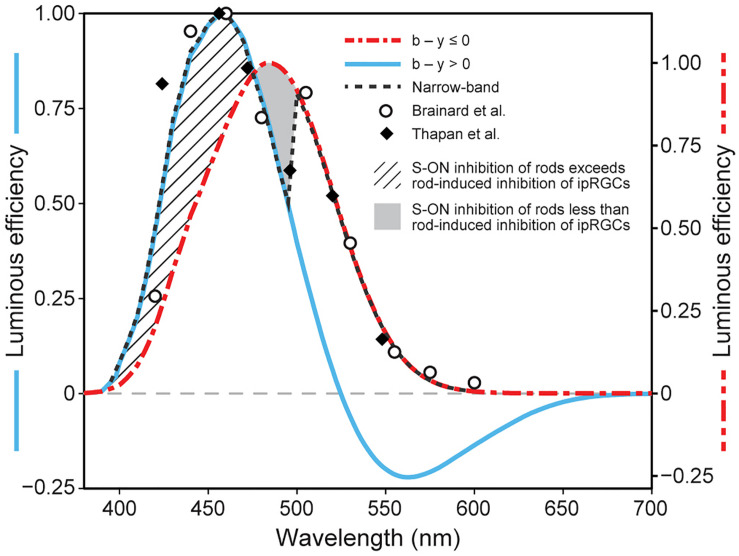
The retina is a complex, but well-organized neural structure that converts optical radiation into neural signals that convey photic information to a wide variety of brain structures. The present paper is concerned with the neural circuits underlying phototransduction for the central pacemaker of the human circadian system. The proposed neural framework adheres to orthodox retinal neuroanatomy and neurophysiology. Several postulated mechanisms are also offered to account for the high threshold and for the subadditive response to polychromatic light exhibited by the human circadian phototransduction circuit. A companion paper, modeling circadian phototransduction: Quantitative predictions of psychophysical data, provides a computational model for predicting psychophysical data associated with nocturnal melatonin suppression while staying within the constraints of the neurophysiology and neuroanatomy offered here.
Keywords: circadian phototransduction; photic sub-additivity; retinal neuroanatomy; retinal neurophysiology; shunting inhibition.
Copyright © 2021 Rea, Nagare and Figueiro.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Modeling Circadian Phototransduction: Quantitative Predictions of Psychophysical Data.Front Neurosci. 2021 Feb 5;15:615322. doi: 10.3389/fnins.2021.615322. eCollection 2021. Front Neurosci. 2021. PMID: 33613181 Free PMC article.
-
Demonstration of additivity failure in human circadian phototransduction.Neuro Endocrinol Lett. 2005 Oct;26(5):493-8. Neuro Endocrinol Lett. 2005. PMID: 16264413 Clinical Trial.
-
Retinal mechanisms determine the subadditive response to polychromatic light by the human circadian system.Neurosci Lett. 2008 Jun 20;438(2):242-5. doi: 10.1016/j.neulet.2008.04.055. Epub 2008 Apr 20. Neurosci Lett. 2008. PMID: 18479818
-
A model of phototransduction by the human circadian system.Brain Res Brain Res Rev. 2005 Dec 15;50(2):213-28. doi: 10.1016/j.brainresrev.2005.07.002. Epub 2005 Oct 7. Brain Res Brain Res Rev. 2005. PMID: 16216333 Review.
-
Circadian organization of the mammalian retina: from gene regulation to physiology and diseases.Prog Retin Eye Res. 2014 Mar;39:58-76. doi: 10.1016/j.preteyeres.2013.12.001. Epub 2013 Dec 12. Prog Retin Eye Res. 2014. PMID: 24333669 Free PMC article. Review.
Cited by
-
Intermittent Light Exposures in Humans: A Case for Dual Entrainment in the Treatment of Alzheimer's Disease.Front Neurol. 2021 Mar 9;12:625698. doi: 10.3389/fneur.2021.625698. eCollection 2021. Front Neurol. 2021. PMID: 33767659 Free PMC article. Review.
-
Individual light history matters to deal with the Antarctic summer.Sci Rep. 2023 Jul 26;13(1):12081. doi: 10.1038/s41598-023-39315-y. Sci Rep. 2023. PMID: 37495664 Free PMC article.
-
Supplemental ambient lighting intervention to improve sleep in Parkinson's disease: A pilot trial.Parkinsonism Relat Disord. 2024 Dec;129:107149. doi: 10.1016/j.parkreldis.2024.107149. Epub 2024 Sep 20. Parkinsonism Relat Disord. 2024. PMID: 39405632 Free PMC article. Clinical Trial.
-
High circadian stimulus lighting therapy for depression: Meta-analysis of clinical trials.Front Neurosci. 2022 Oct 6;16:975576. doi: 10.3389/fnins.2022.975576. eCollection 2022. Front Neurosci. 2022. PMID: 36278019 Free PMC article.
-
The circadian stimulus-oscillator model: Improvements to Kronauer's model of the human circadian pacemaker.Front Neurosci. 2022 Sep 27;16:965525. doi: 10.3389/fnins.2022.965525. eCollection 2022. Front Neurosci. 2022. PMID: 36238087 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources