

Metabolic Potential for Reductive Acetogenesis and a Novel Energy-Converting [NiFe] Hydrogenase in Bathyarchaeia From Termite Guts - A Genome-Centric Analysis
- PMID: 33613473
- PMCID: PMC7886697
- DOI: 10.3389/fmicb.2020.635786
Metabolic Potential for Reductive Acetogenesis and a Novel Energy-Converting [NiFe] Hydrogenase in Bathyarchaeia From Termite Guts - A Genome-Centric Analysis
Abstract
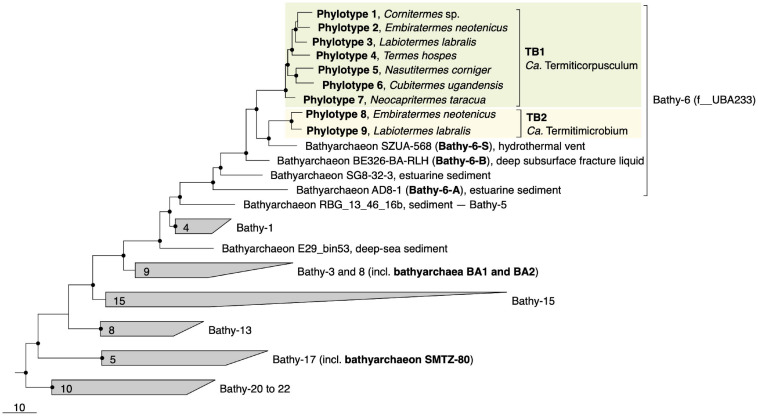
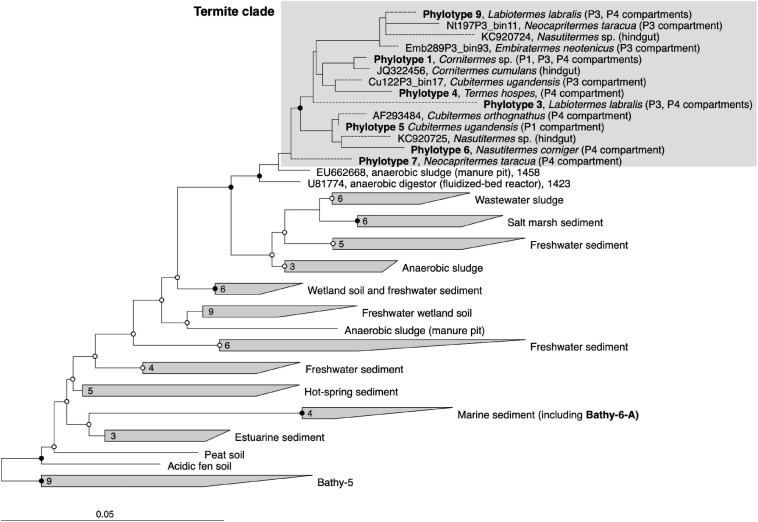
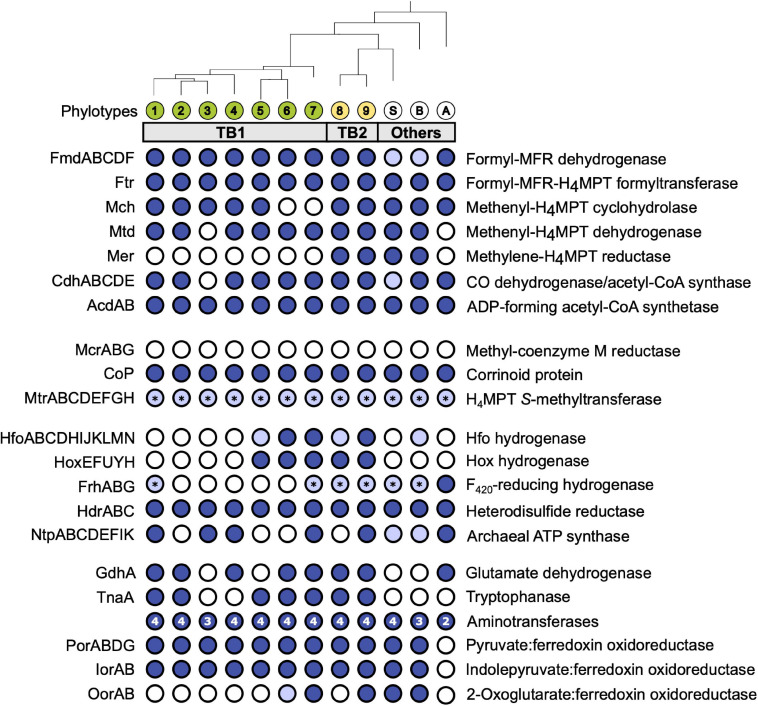
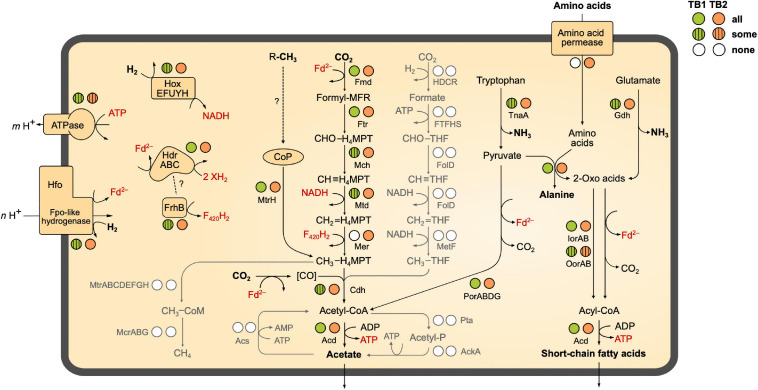
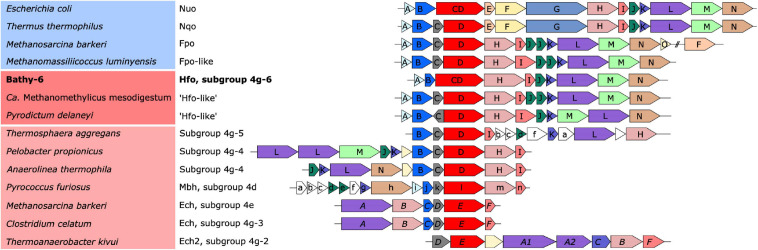
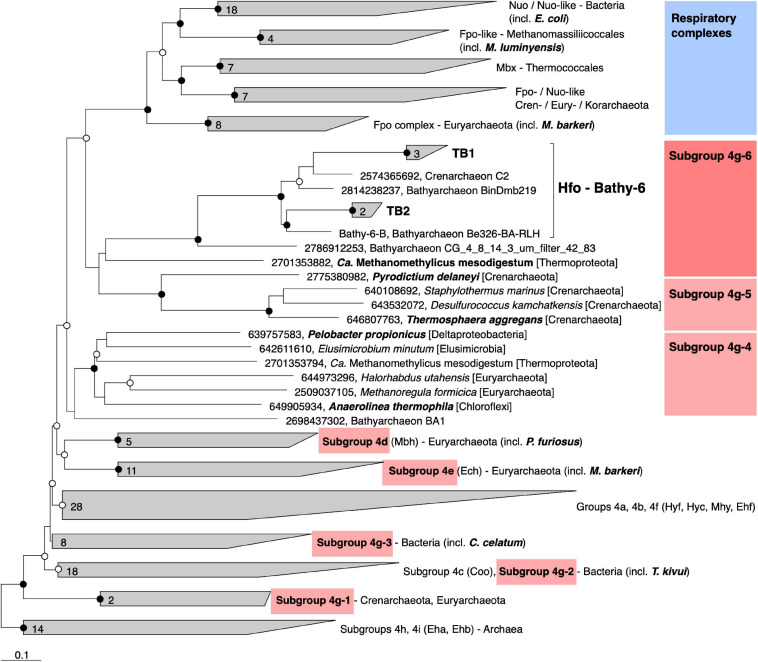
Symbiotic digestion of lignocellulose in the hindgut of higher termites is mediated by a diverse assemblage of bacteria and archaea. During a large-scale metagenomic study, we reconstructed 15 metagenome-assembled genomes of Bathyarchaeia that represent two distinct lineages in subgroup 6 (formerly MCG-6) unique to termite guts. One lineage (TB2; Candidatus Termitimicrobium) encodes all enzymes required for reductive acetogenesis from CO2 via an archaeal variant of the Wood-Ljungdahl pathway, involving tetrahydromethanopterin as C1 carrier and an (ADP-forming) acetyl-CoA synthase. This includes a novel 11-subunit hydrogenase, which possesses the genomic architecture of the respiratory Fpo-complex of other archaea but whose catalytic subunit is phylogenetically related to and shares the conserved [NiFe] cofactor-binding motif with [NiFe] hydrogenases of subgroup 4 g. We propose that this novel Fpo-like hydrogenase provides part of the reduced ferredoxin required for CO2 reduction and is driven by the electrochemical membrane potential generated from the ATP conserved by substrate-level phosphorylation; the other part may require the oxidation of organic electron donors, which would make members of TB2 mixotrophic acetogens. Members of the other lineage (TB1; Candidatus Termiticorpusculum) are definitely organotrophic because they consistently lack hydrogenases and/or methylene-tetrahydromethanopterin reductase, a key enzyme of the archaeal Wood-Ljungdahl pathway. Both lineages have the genomic capacity to reduce ferredoxin by oxidizing amino acids and might conduct methylotrophic acetogenesis using unidentified methylated compound(s). Our results indicate that Bathyarchaeia of subgroup 6 contribute to acetate formation in the guts of higher termites and substantiate the genomic evidence for reductive acetogenesis from organic substrates, possibly including methylated compounds, in other uncultured representatives of the phylum.
Keywords: Bathyarchaeota; Wood-Ljungdahl pathway; acetogens; comparative genomics; gut microbiota; metagenome-assembled genomes; termites.
Copyright © 2021 Loh, Hervé and Brune.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

Similar articles
-
Genomic and in-situ Transcriptomic Characterization of the Candidate Phylum NPL-UPL2 From Highly Alkaline Highly Reducing Serpentinized Groundwater.Front Microbiol. 2018 Dec 18;9:3141. doi: 10.3389/fmicb.2018.03141. eCollection 2018. Front Microbiol. 2018. PMID: 30619209 Free PMC article.
-
Genomic Analysis of Calderihabitans maritimus KKC1, a Thermophilic, Hydrogenogenic, Carboxydotrophic Bacterium Isolated from Marine Sediment.Appl Environ Microbiol. 2017 Jul 17;83(15):e00832-17. doi: 10.1128/AEM.00832-17. Print 2017 Aug 1. Appl Environ Microbiol. 2017. PMID: 28526793 Free PMC article.
-
The Polar Fox Lagoon in Siberia harbours a community of Bathyarchaeota possessing the potential for peptide fermentation and acetogenesis.Antonie Van Leeuwenhoek. 2022 Oct;115(10):1229-1244. doi: 10.1007/s10482-022-01767-z. Epub 2022 Aug 10. Antonie Van Leeuwenhoek. 2022. PMID: 35947314 Free PMC article.
-
Hydrogenases from methanogenic archaea, nickel, a novel cofactor, and H2 storage.Annu Rev Biochem. 2010;79:507-36. doi: 10.1146/annurev.biochem.030508.152103. Annu Rev Biochem. 2010. PMID: 20235826 Review.
-
Energy-converting [NiFe] hydrogenases from archaea and extremophiles: ancestors of complex I.J Bioenerg Biomembr. 2004 Feb;36(1):65-75. doi: 10.1023/b:jobb.0000019599.43969.33. J Bioenerg Biomembr. 2004. PMID: 15168611 Review.
Cited by
-
Functional insights of novel Bathyarchaeia reveal metabolic versatility in their role in peatlands of the Peruvian Amazon.Microbiol Spectr. 2024 Nov 14;12(12):e0038724. doi: 10.1128/spectrum.00387-24. Online ahead of print. Microbiol Spectr. 2024. PMID: 39540749 Free PMC article.
-
Genomic remnants of ancestral methanogenesis and hydrogenotrophy in Archaea drive anaerobic carbon cycling.Sci Adv. 2022 Nov 4;8(44):eabm9651. doi: 10.1126/sciadv.abm9651. Epub 2022 Nov 4. Sci Adv. 2022. PMID: 36332026 Free PMC article.
-
Factors shaping the abundance and diversity of the gut archaeome across the animal kingdom.Nat Commun. 2022 Jun 10;13(1):3358. doi: 10.1038/s41467-022-31038-4. Nat Commun. 2022. PMID: 35688919 Free PMC article.
-
Metabolic features that select for Bathyarchaeia in modern ferruginous lacustrine subsurface sediments.ISME Commun. 2024 Sep 14;4(1):ycae112. doi: 10.1093/ismeco/ycae112. eCollection 2024 Jan. ISME Commun. 2024. PMID: 39660009 Free PMC article.
-
Insights into Bacterial Communities and Diversity of Mangrove Forest Soils along the Upper Gulf of Thailand in Response to Environmental Factors.Biology (Basel). 2022 Dec 8;11(12):1787. doi: 10.3390/biology11121787. Biology (Basel). 2022. PMID: 36552296 Free PMC article.
References
-
- Altschul S. F., Gish W., Miller W., Myers E. W., Lipman D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215 403–410. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources