

Immune Evasion Strategies of Schistosomes
- PMID: 33613562
- PMCID: PMC7889519
- DOI: 10.3389/fimmu.2020.624178
Immune Evasion Strategies of Schistosomes
Abstract
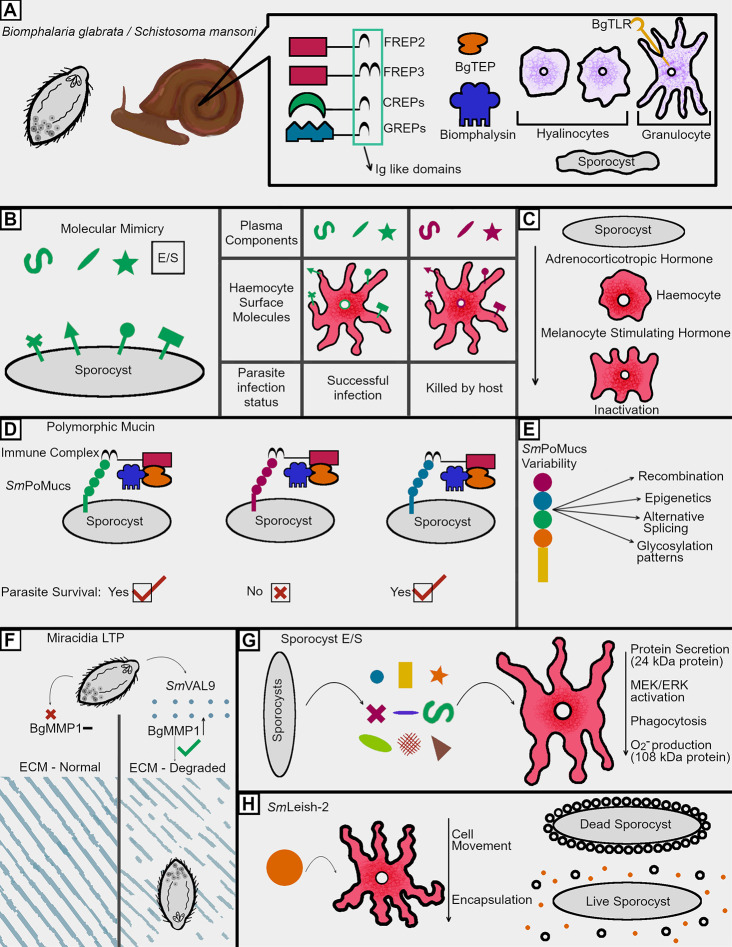
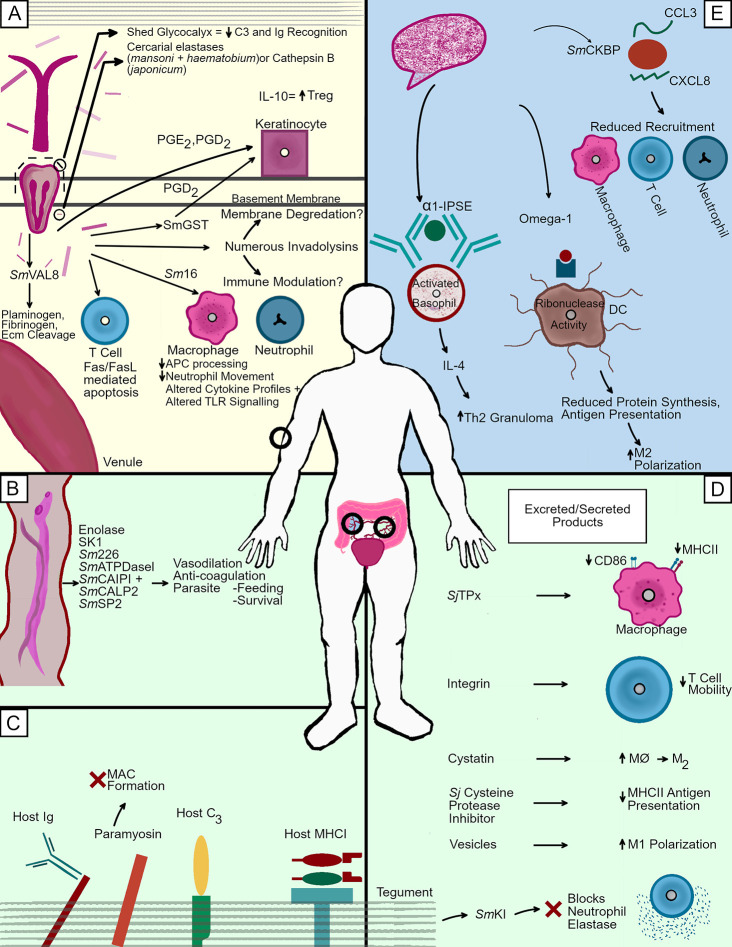
Human schistosomes combat the unique immune systems of two vastly different hosts during their indirect life cycles. In gastropod molluscs, they face a potent innate immune response composed of variable immune recognition molecules and highly phagocytic hemocytes. In humans, a wide variety of innate and adaptive immune processes exist in proximity to these parasites throughout their lifespan. To survive and thrive as the second most common parasitic disease in humans, schistosomes have evolved many techniques to avoid and combat these targeted host responses. Among these techniques are molecular mimicry of host antigens, the utilization of an immune resistant outer tegument, the secretion of several potent proteases, and targeted release of specific immunomodulatory factors affecting immune cell functions. This review seeks to describe these key immune evasion mechanisms, among others, which schistosomes use to survive in both of their hosts. After diving into foundational observational studies of the processes mediating the establishment of schistosome infections, more recent transcriptomic and proteomic studies revealing crucial components of the host/parasite molecular interface are discussed. In order to combat this debilitating and lethal disease, a comprehensive understanding of schistosome immune evasion strategies is necessary for the development of novel therapeutics and treatment plans, necessitating the discussion of the numerous ways in which these parasitic flatworms overcome the immune responses of both hosts.
Keywords: Biomphalaria glabrata; immune evasion; immunomodulation; schistosome; schistosomiasis.
Copyright © 2021 Hambrook and Hanington.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Ezeamama AE, Bustinduy AL, Nkwata AK, Martinez L, Pabalan N, Boivin MJ, et al. Cognitive deficits and educational loss in children with schistosome infection—A systematic review and meta-analysis. PLoS Negl Trop Dis (2018) 12(1):e0005524. 10.1371/journal.pntd.0005524. Garba A, editor. - DOI - PMC - PubMed
-
- Global Health, Division of Parasitic Diseases and Malaria CDC - Global Health - Neglected Tropical Diseases. Centers for Disease Control and Prevention (2020). Available at: https://www.cdc.gov/globalhealth/ntd/index.html
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources