

Engineering highly efficient backsplicing and translation of synthetic circRNAs
- PMID: 33614232
- PMCID: PMC7868716
- DOI: 10.1016/j.omtn.2021.01.003
Engineering highly efficient backsplicing and translation of synthetic circRNAs
Abstract
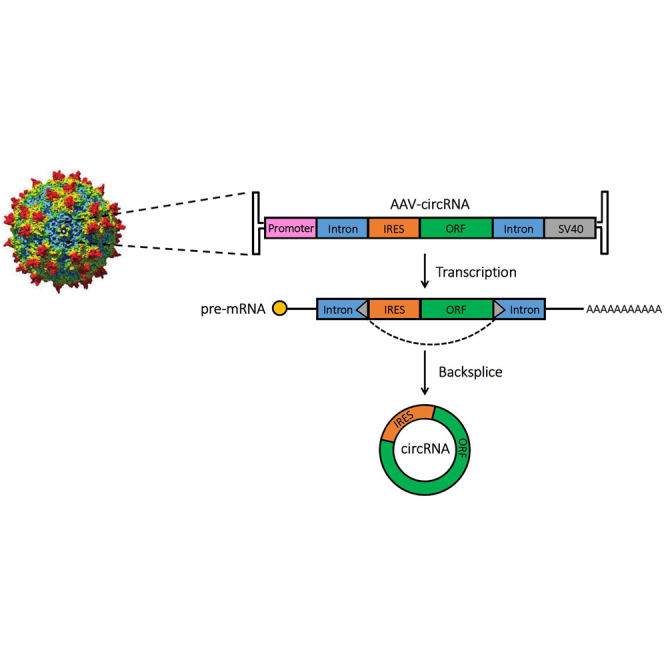
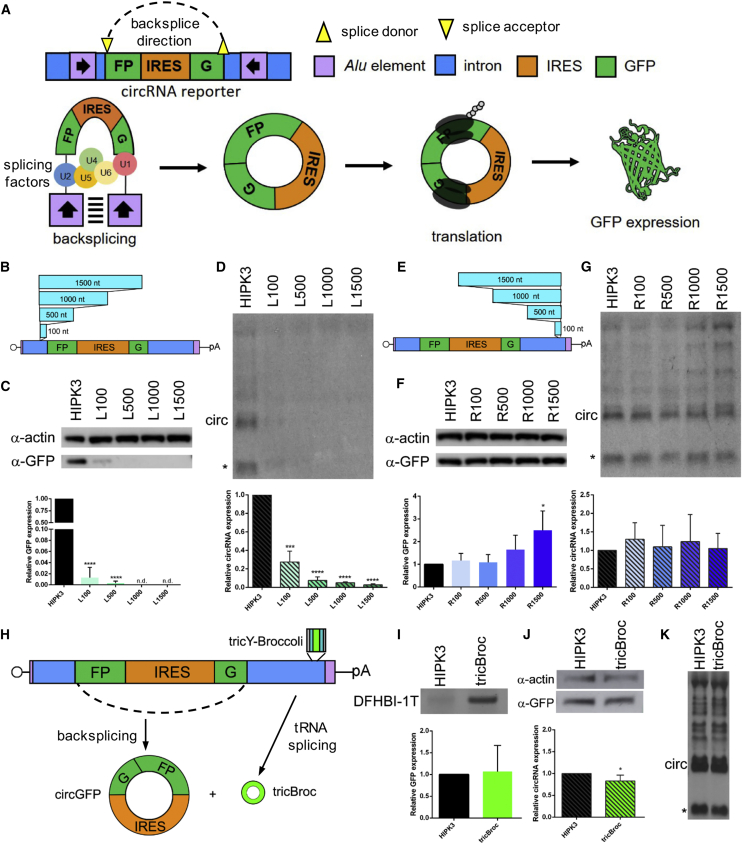
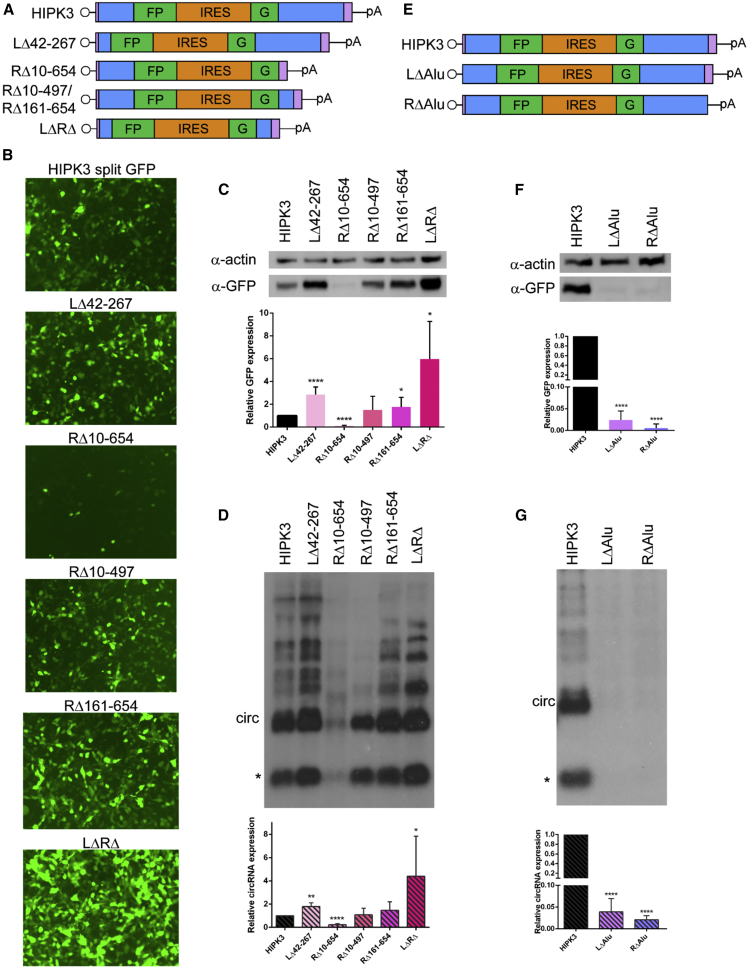
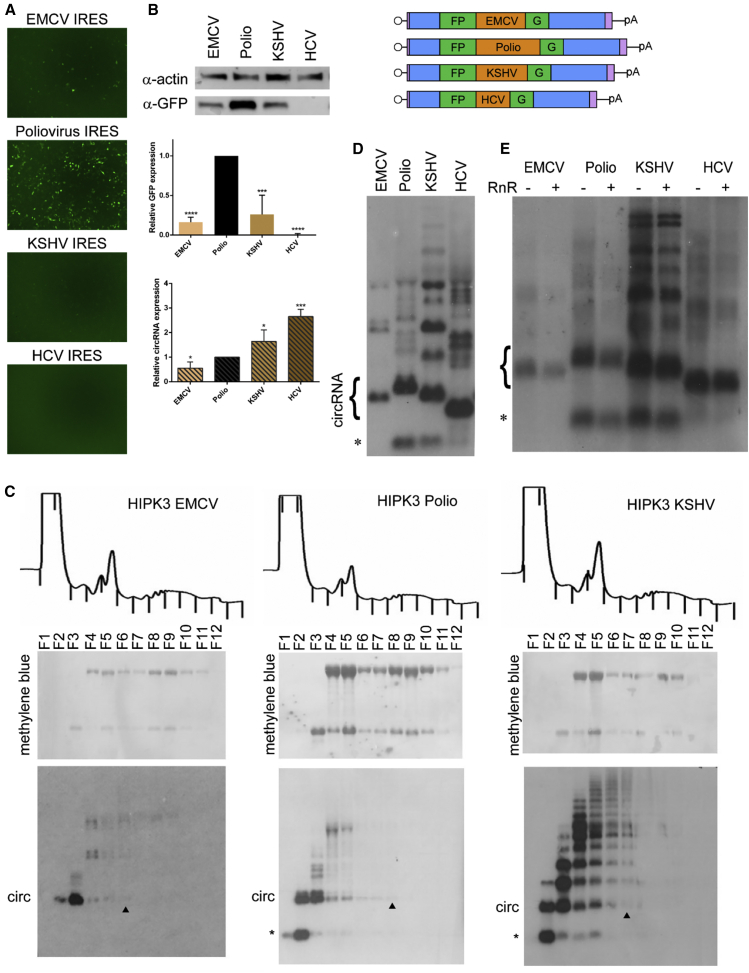
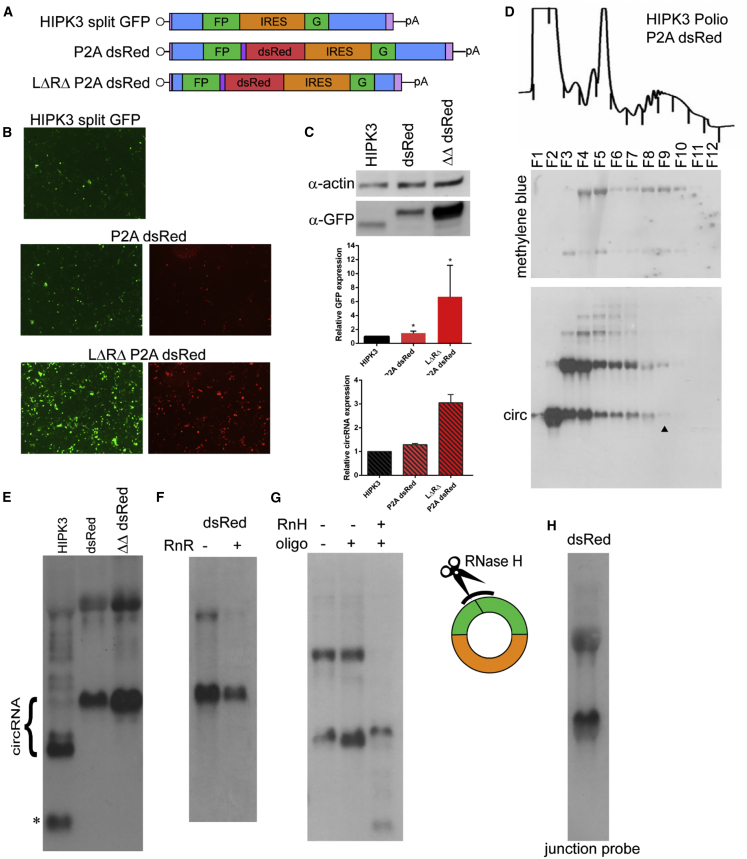
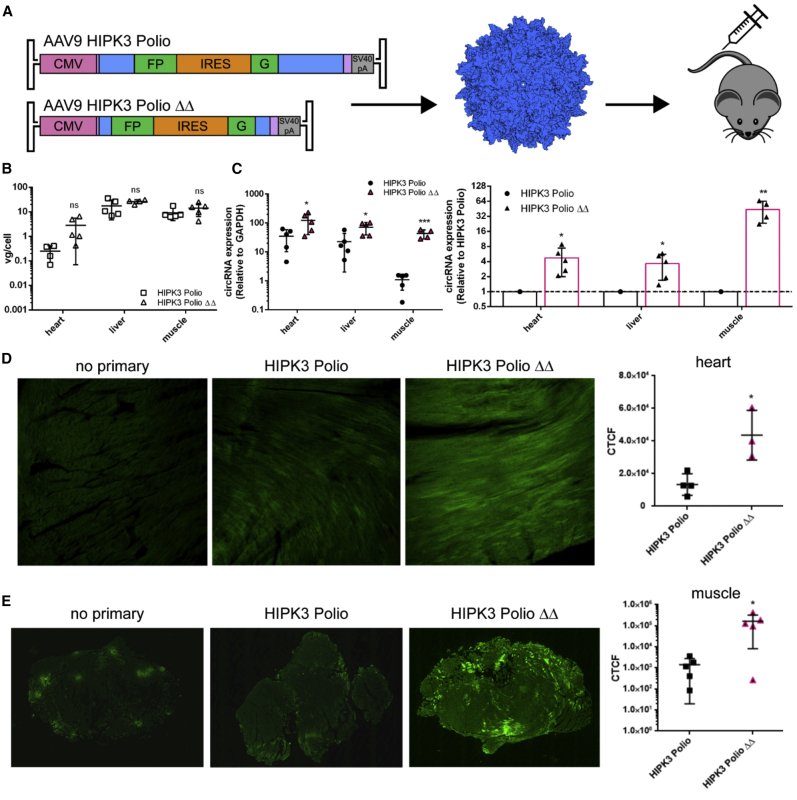
Circular RNAs (circRNAs) are highly stable RNA molecules that are attractive templates for expression of therapeutic proteins and non-coding RNAs. In eukaryotes, circRNAs are primarily generated by the spliceosome through backsplicing. Here, we interrogate different molecular elements including intron type and length, Alu repeats, internal ribosome entry sites (IRESs), and exon length essential for circRNA formation and exploit this information to engineer robust backsplicing and circRNA expression. Specifically, we leverage the finding that the downstream intron can tolerate large inserts without affecting splicing to achieve tandem expression of backspliced circRNAs and tRNA intronic circRNAs from the same template. Further, truncation of selected intronic regions markedly increased circRNA formation in different cell types in vitro as well as AAV-mediated circRNA expression in cardiac and skeletal muscle tissue in vivo. We also observed that different IRES elements and exon length influenced circRNA expression and translation, revealing an exonic contribution to splicing, as evidenced by different RNA species produced. Taken together, these data provide new insight into improving the design and expression of synthetic circRNAs. When combined with AAV capsid and promoter technologies, the backsplicing introns and IRES elements constituting this modular platform significantly expand the gene expression toolkit.
© 2021 The Authors.
Conflict of interest statement
A.A., W.F.M, and R.M.M. are inventors on patent applications filed on the subject matter of this manuscript. A.A. is a co-founder at TorqueBio, LLC.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
