

Compartmentalization of Photoreceptor Sensory Cilia
- PMID: 33614665
- PMCID: PMC7889997
- DOI: 10.3389/fcell.2021.636737
Compartmentalization of Photoreceptor Sensory Cilia
Abstract
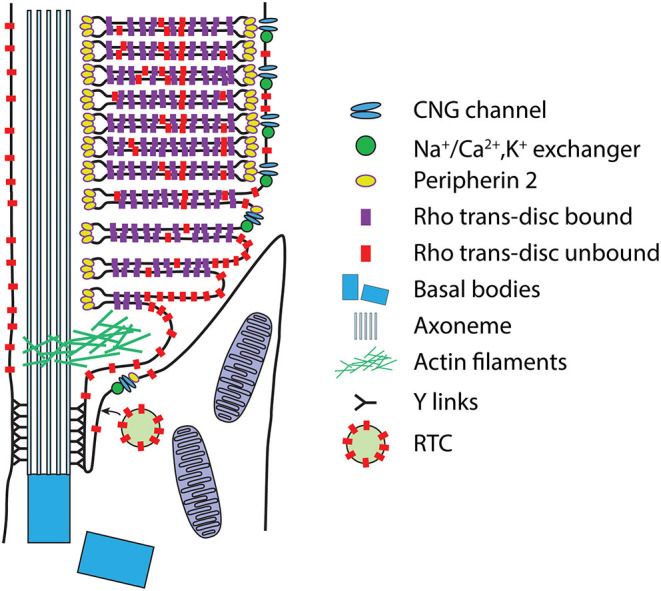
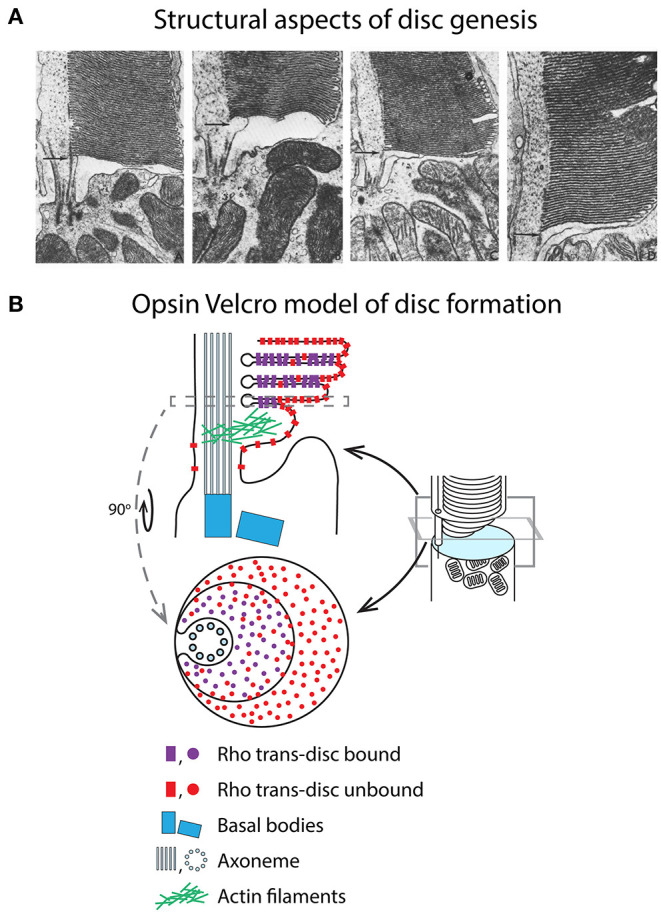
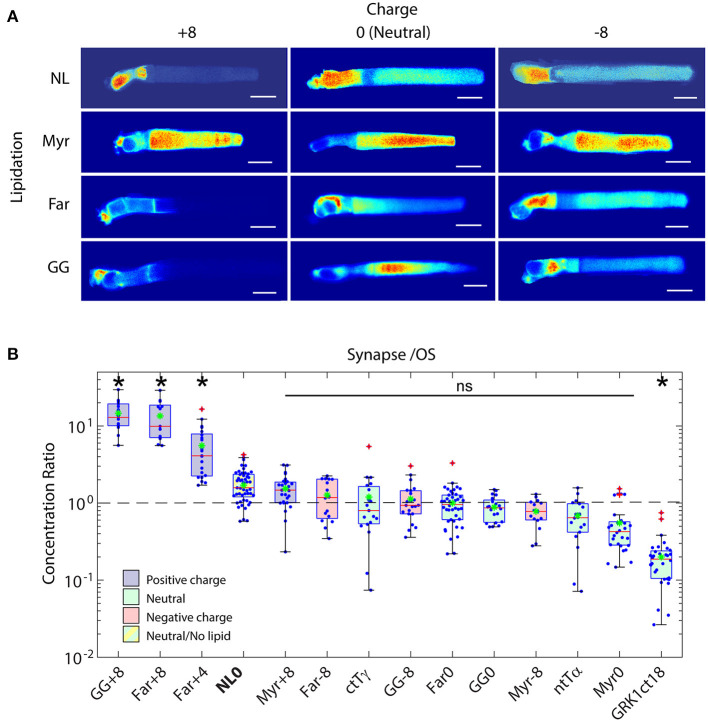
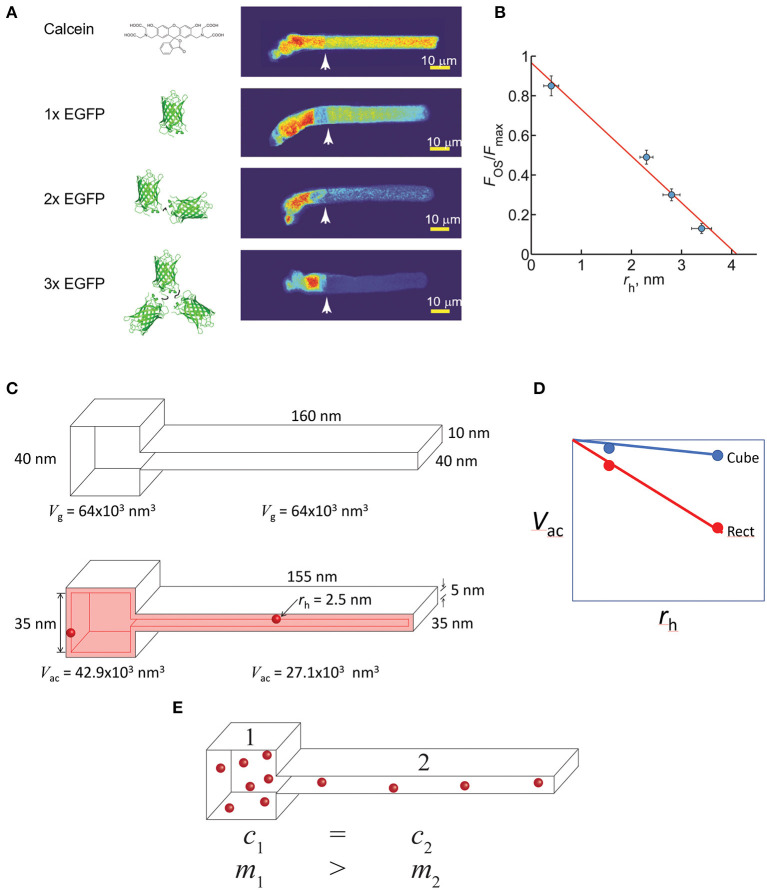
Functional compartmentalization of cells is a universal strategy for segregating processes that require specific components, undergo regulation by modulating concentrations of those components, or that would be detrimental to other processes. Primary cilia are hair-like organelles that project from the apical plasma membranes of epithelial cells where they serve as exclusive compartments for sensing physical and chemical signals in the environment. As such, molecules involved in signal transduction are enriched within cilia and regulating their ciliary concentrations allows adaptation to the environmental stimuli. The highly efficient organization of primary cilia has been co-opted by major sensory neurons, olfactory cells and the photoreceptor neurons that underlie vision. The mechanisms underlying compartmentalization of cilia are an area of intense current research. Recent findings have revealed similarities and differences in molecular mechanisms of ciliary protein enrichment and its regulation among primary cilia and sensory cilia. Here we discuss the physiological demands on photoreceptors that have driven their evolution into neurons that rely on a highly specialized cilium for signaling changes in light intensity. We explore what is known and what is not known about how that specialization appears to have driven unique mechanisms for photoreceptor protein and membrane compartmentalization.
Keywords: outer segment; peripheral membrane protein; photoreceptors; primary cilia; rhodopsin; soluble protein; trafficking; transport.
Copyright © 2021 Barnes, Malhotra and Calvert.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
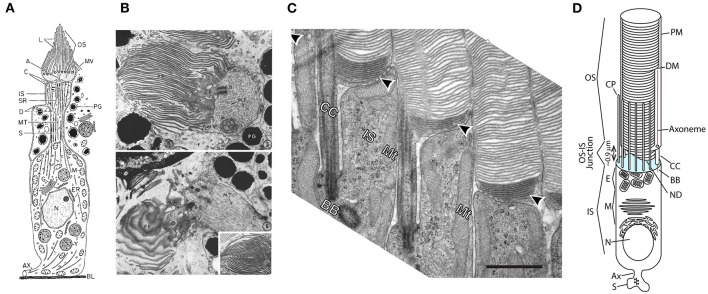
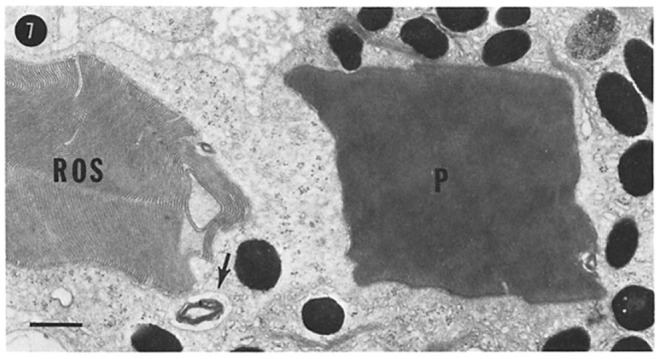
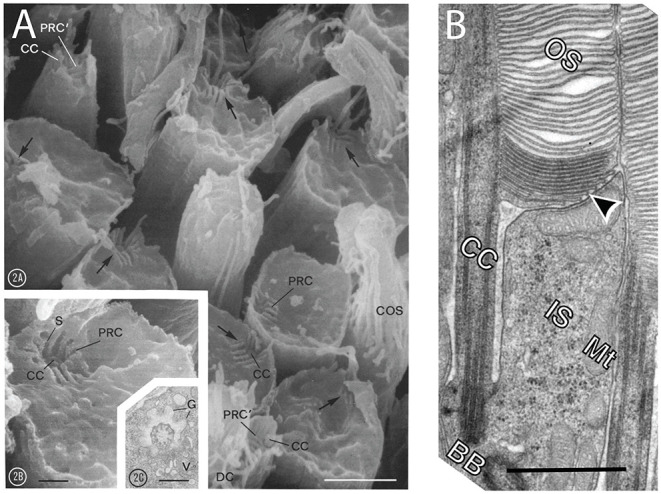
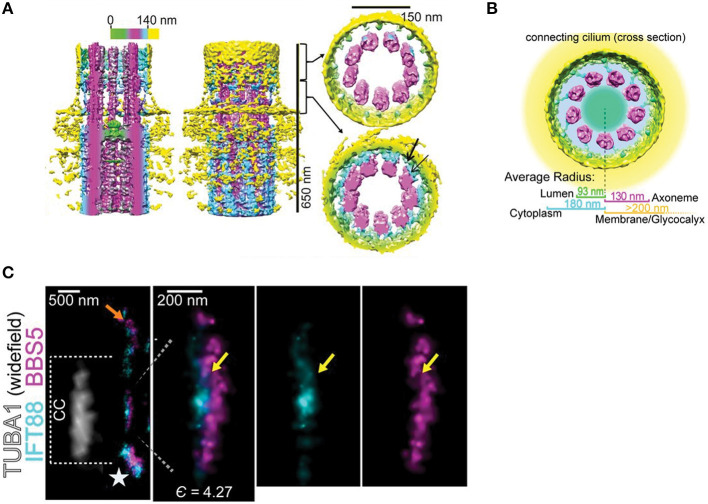
References
-
- Alpadi K., Magupalli V. G., Käppel S., Köblitz L., Schwarz K., Seigel G. M., et al. . (2008). RIBEYE recruits Munc119, a mammalian ortholog of the Caenorhabditis elegans protein unc119, to synaptic ribbons of photoreceptor synapses. J. Biol. Chem. 283, 26461–26467. 10.1074/jbc.M801625200 - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials