

The mitochondrial AAA protease FTSH3 regulates Complex I abundance by promoting its disassembly
- PMID: 33616659
- PMCID: PMC8154063
- DOI: 10.1093/plphys/kiab074
The mitochondrial AAA protease FTSH3 regulates Complex I abundance by promoting its disassembly
Abstract
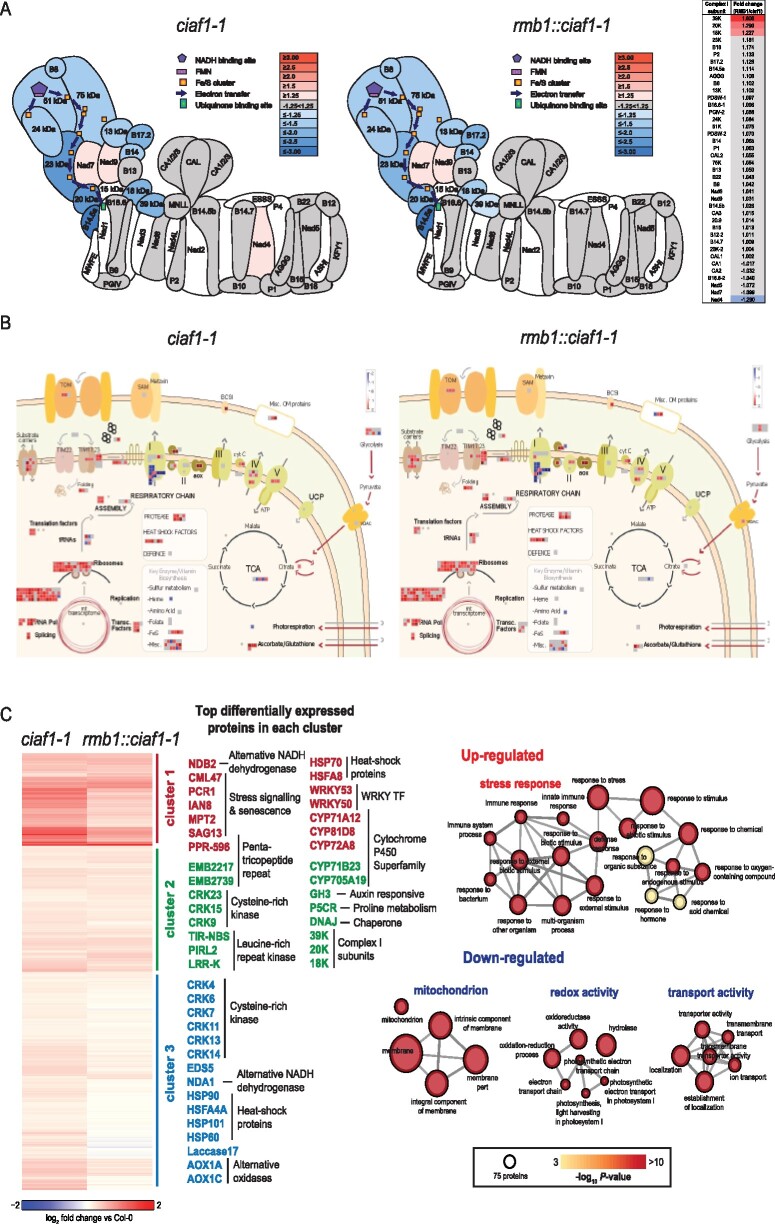
ATP is generated in mitochondria by oxidative phosphorylation. Complex I (NADH:ubiquinone oxidoreductase or NADH dehydrogenase) is the first multisubunit protein complex of this pathway, oxidizing NADH and transferring electrons to the ubiquinone pool. Typically, Complex I mutants display a slow growth rate compared to wild-type plants. Here, using a forward genetic screen approach for restored growth of a Complex I mutant, we have identified the mitochondrial ATP-dependent metalloprotease, Filamentous Temperature Sensitive H 3 (FTSH3), as a factor that is required for the disassembly of Complex I. An ethyl methanesulfonate-induced mutation in FTSH3, named as rmb1 (restoration of mitochondrial biogenesis 1), restored Complex I abundance and plant growth. Complementation could be achieved with FTSH3 lacking proteolytic activity, suggesting the unfoldase function of FTSH3 has a role in Complex I disassembly. The introduction of the rmb1 to an additional, independent, and extensively characterized Complex I mutant, ndufs4, resulted in similar increases to Complex I abundance and a partial restoration of growth. These results show that disassembly or degradation of Complex I plays a role in determining its steady-state abundance and thus turnover may vary under different conditions.
© American Society of Plant Biologists 2021. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures




References
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al. (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
