

Convergent antibody evolution and clonotype expansion following influenza virus vaccination
- PMID: 33617543
- PMCID: PMC7899375
- DOI: 10.1371/journal.pone.0247253
Convergent antibody evolution and clonotype expansion following influenza virus vaccination
Abstract
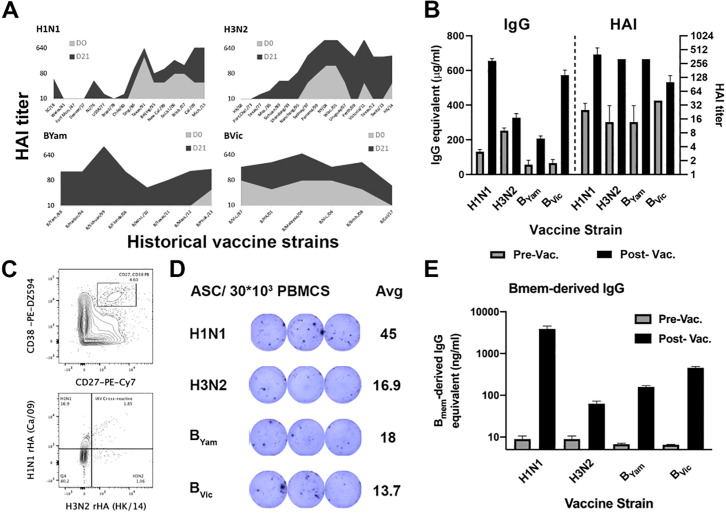
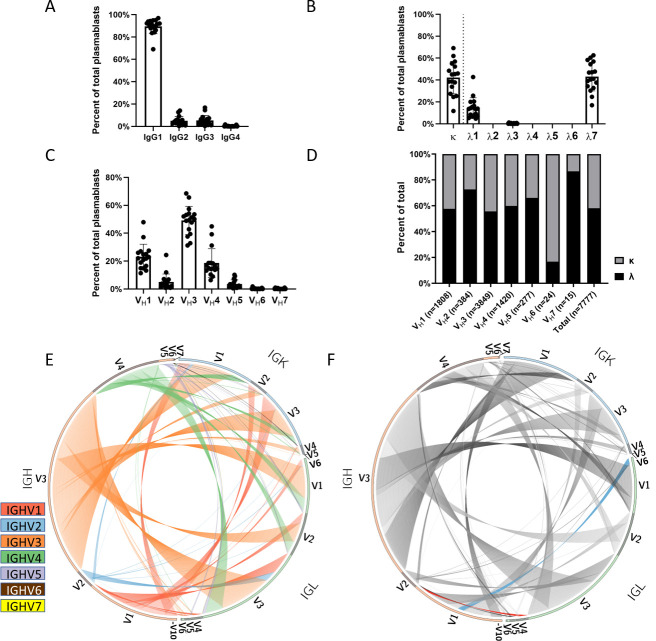
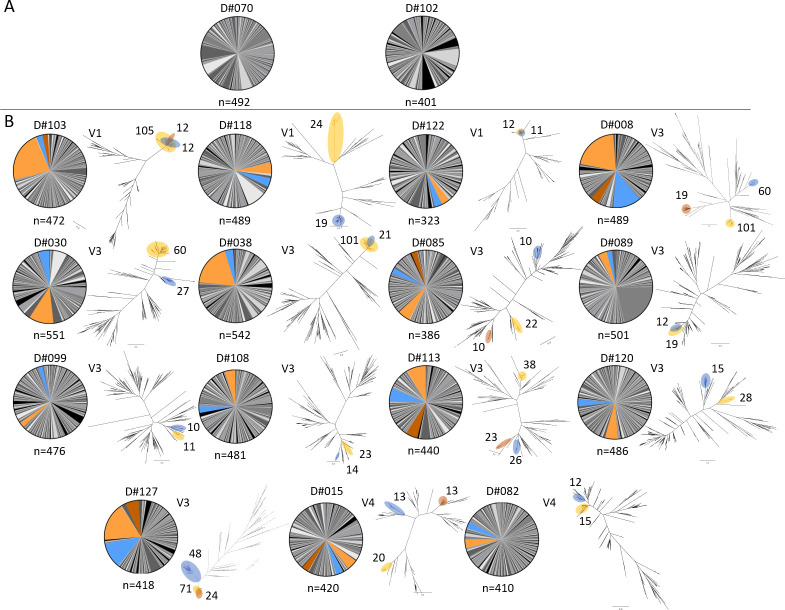
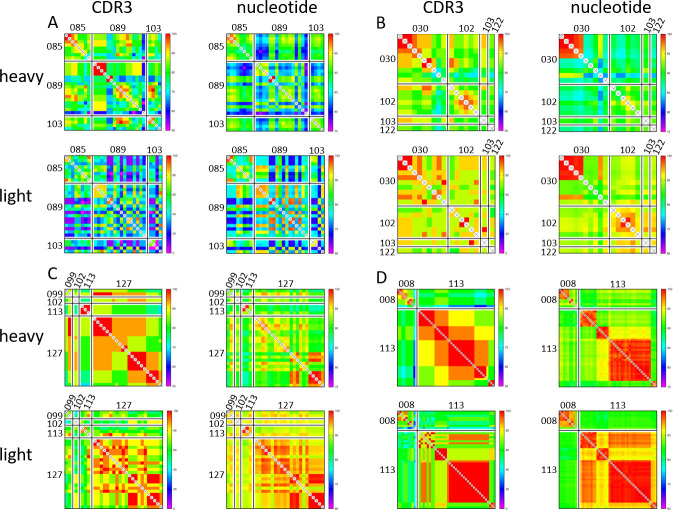
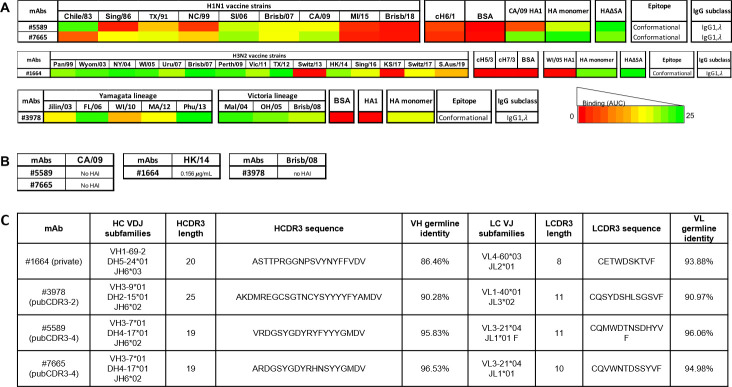
Recent advances in high-throughput single cell sequencing have opened up new avenues into the investigation of B cell receptor (BCR) repertoires. In this study, PBMCs were collected from 17 human participants vaccinated with the split-inactivated influenza virus vaccine during the 2016-2017 influenza season. A combination of Immune Repertoire Capture (IRCTM) technology and IgG sequencing was performed on ~7,800 plasmablast (PB) cells and preferential IgG heavy-light chain pairings were investigated. In some participants, a single expanded clonotype accounted for ~22% of their PB BCR repertoire. Approximately 60% (10/17) of participants experienced convergent evolution, possessing public PBs that were elicited independently in multiple participants. Binding profiles of one private and three public PBs confirmed they were all subtype-specific, cross-reactive hemagglutinin (HA) head-directed antibodies. Collectively, this high-resolution antibody repertoire analysis demonstrated the impact evolution can have on BCRs in response to influenza virus vaccination, which can guide future universal influenza prophylactic approaches.
Conflict of interest statement
Some of the authors are affiliated with Atreca, Inc., but the company provided no funding for the study and no competing interests exist. The commercial affiliation of those authors does not alter our adherence to PLoS ONE policies on sharing data and materials.
Figures

References
-
- Francis ME, McNeil M, Dawe NJ, Foley MK, King ML, Ross TM, et al. Historical H1N1 influenza virus imprinting increases vaccine protection by influencing the activity and sustained production of antibodies elicited at vaccination in ferrets. Vaccines. 2019;7: 133 10.3390/vaccines7040133 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
