

Unfolded and intermediate states of PrP play a key role in the mechanism of action of an antiprion chaperone
- PMID: 33619087
- PMCID: PMC7936342
- DOI: 10.1073/pnas.2010213118
Unfolded and intermediate states of PrP play a key role in the mechanism of action of an antiprion chaperone
Abstract
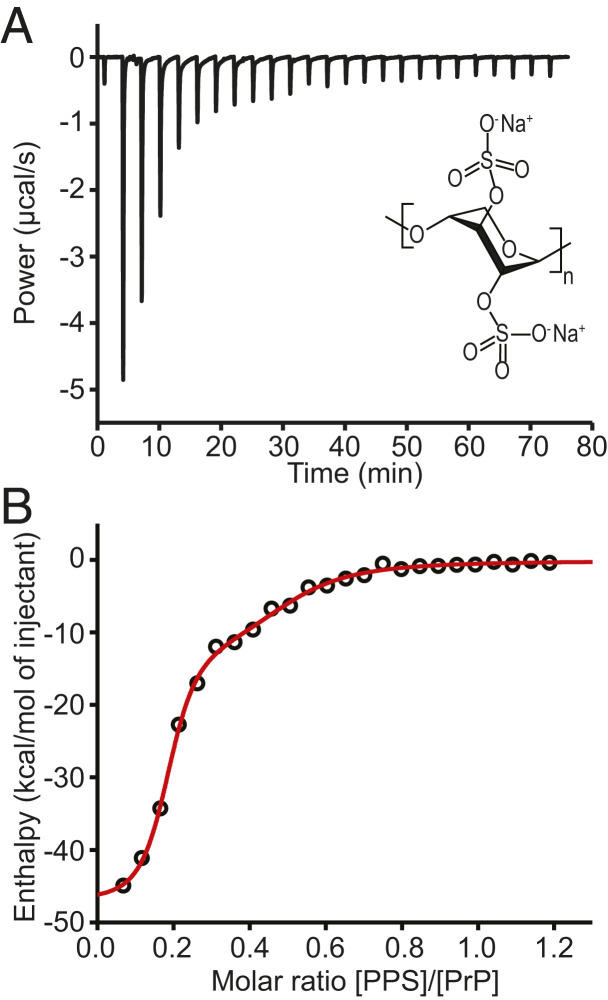
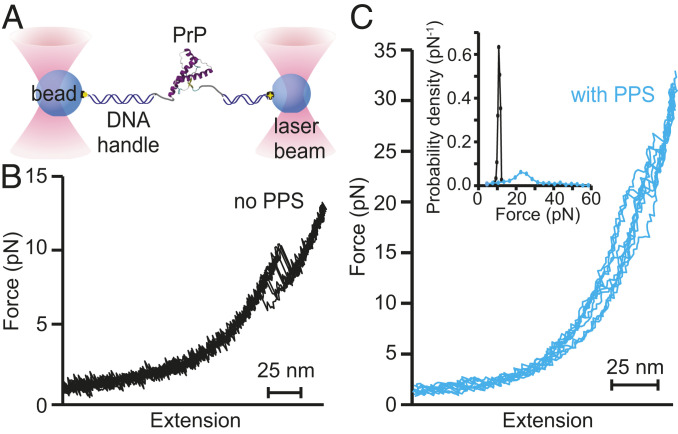
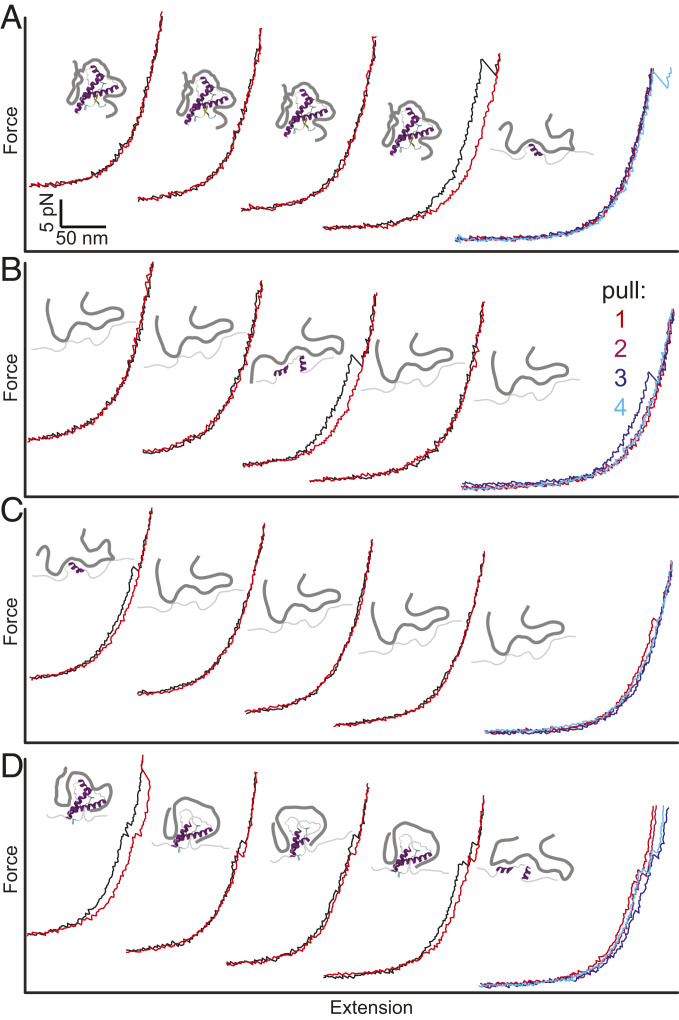
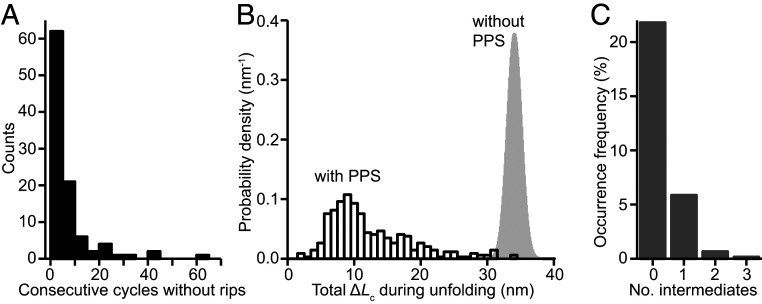
Prion and prion-like diseases involve the propagation of misfolded protein conformers. Small-molecule pharmacological chaperones can inhibit propagated misfolding, but how they interact with disease-related proteins to prevent misfolding is often unclear. We investigated how pentosan polysulfate (PPS), a polyanion with antiprion activity in vitro and in vivo, interacts with mammalian prion protein (PrP) to alter its folding. Calorimetry showed that PPS binds two sites on natively folded PrP, but one PPS molecule can bind multiple PrP molecules. Force spectroscopy measurements of single PrP molecules showed PPS stabilizes not only the native fold of PrP but also many different partially folded intermediates that are not observed in the absence of PPS. PPS also bound tightly to unfolded segments of PrP, delaying refolding. These observations imply that PPS can act through multiple possible modes, inhibiting misfolding not only by stabilizing the native fold or sequestering natively folded PrP into aggregates, as proposed previously, but also by binding to partially or fully unfolded states that play key roles in mediating misfolding. These results underline the likely importance of unfolded states as critical intermediates on the prion conversion pathway.
Keywords: energy landscape; optical tweezers; pharmacological chaperone; protein misfolding.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Chiti F., Dobson C. M., Protein misfolding, amyloid formation, and human disease: A summary of progress over the last decade. Annu. Rev. Biochem. 86, 27–68 (2017). - PubMed
-
- Hartl F. U., Bracher A., Hayer-Hartl M., Molecular chaperones in protein folding and proteostasis. Nature 475, 324–332 (2011). - PubMed
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
